definisi replikasi DNA. proses replikasi DNA. Tiga prinsip struktur DNA
Divisi sel terjadi melalui mitosis, sementara, untuk menghindari hilangnya informasi genetik, seluruh genom nuklir pertama kali berlipat ganda dalam fase-S dari siklus sel. Durasi fase S adalah 8 jam. DNA sentromer kromosom bereplikasi selama tahap tengah mitosis, yang mendahului proses segregasi kromosom.
replikasi mitokondria dan nukleus terjadi pada fase yang berbeda dari siklus sel. Terlepas dari kenyataan bahwa urutan umum tahapan dalam replikasi DNA inti pada makhluk yang lebih tinggi (eukariota) dan bakteri (prokariota) adalah sama, prosesnya sendiri memiliki perbedaan kecil. Jadi, pada eukariota, selama replikasi, DNA (nuklir) tetap dalam konfigurasi nukleosom.
Fragmen DNA, kaya akan pasangan basa G-C (pita-R eukromatin dalam kromatin yang dipadatkan), mengekspresikan gen rumah tangga yang berfungsi di semua sel tubuh. Fragmen ini direplikasi di awal fase S. Daerah heterokromatin yang kaya akan pasangan basa A-T (G-band) mengekspresikan sejumlah kecil gen dan bereplikasi di akhir fase S.
gen dengan kandungan pasangan A-T yang tinggi, mengkodekan berbagai sifat dan hanya berfungsi dalam sel tertentu, merupakan bagian dari heterokromatin fakultatif. Replikasi mereka terjadi pada tahap awal fase-S hanya di sel-sel di mana mereka diekspresikan, dan pada tahap selanjutnya - dalam sel di mana ekspresi tidak terjadi.
Daerah spiral DNA, yang pertama kali melepas pada awal replikasi, disebut situs awal replikasi (replika). Pada titik ini, untai ganda dilepaskan oleh enzim helikase, yang mengungkapkan urutan basa. Proses replikasi terjadi di sepanjang satu untai dengan kecepatan sekitar 40-50 nukleotida per detik secara bersamaan di kedua arah. Makhluk yang lebih tinggi memiliki banyak replika yang terletak pada jarak 50.000-300.000 bp. Di tempat pemisahan untai DNA, pembengkakan replikasi muncul, di setiap ujungnya terbentuk garpu replikasi.
Baru DNA disintesis dengan partisipasi enzim yang disebut DNA polimerase dari deoksiribonukleotida trifosfat (ATP, GTP, dll.), Yang diubah menjadi nukleotida monofosfat (AMP, GMP, dll.). Pembelahan dan hidrolisis pirofosfat dari trifosfat menyediakan proses dengan energi dan membuatnya benar-benar ireversibel, membuat molekul DNA cukup stabil.
Semuanya DNA polimerase dapat membangun DNA baru hanya dalam arah dari ujung 5" ke ujung 3". Ini berarti bahwa enzim harus bergerak sepanjang rantai cetakan dari ujung 3' ke ujung 5'. Dalam hal ini, replikasi dapat terus menerus terjadi dari replika sepanjang hanya satu rantai, yang disebut memajukan. Karena lokasi gula, replikasi sepanjang untai kedua yang tertinggal hanya terjadi pada bentangan pendek yang dikenal sebagai fragmen Okazaki.
Panjang baru fragmen DNA, terbentuk di sepanjang rantai tertinggal, rata-rata 100-200 pasangan basa. Selama sintesis, fragmen Okazaki diikat silang oleh enzim DNA ligase. Untuk mengantisipasi replikasi, stabilitas urutan nukleotida untai tunggal primer dari untai tertinggal dipertahankan oleh protein pengikat DNA untai tunggal (atau protein destabilisasi heliks).
Untuk sintesis terkemuka rantai membutuhkan enzim DNA polimerase S, dan untuk sintesis DNA polimerase tertinggal a. Yang terakhir memiliki subunit yang disebut DNA primase, yang mensintesis primer RNA pendek yang bertindak sebagai primer. Replikasi DNA mitokondria terjadi secara independen dari proses dalam nukleus. Dalam hal ini, sejumlah enzim lain digunakan, salah satunya adalah DNA polimerase y.
Genom berisi sejumlah besar salinan lima gen histon, menghasilkan sintesis banyak histon (terutama selama fase S), yang segera setelah replikasi, mengikat untai DNA baru.
Perlu diperhatikan bahwa proses replikasi disebut semi-konservatif, karena komposisi molekul DNA anak mencakup satu untai primer dan satu untai sintesis.
Replikasi telomer DNA
Masalah utama sintesis DNA di ujung lagging strand adalah bahwa DNA polimerase a perlu menempel di atas ujung urutan yang direplikasi dan bekerja secara proksimal dalam arah dari ujung 5" ke ujung 3". Untuk mengatasi masalah ini, diperlukan enzim sintetik DNA, telomerase, yang memperpanjang rantai lagging.
Telomerase- ribonukleoprotein yang mengandung RNA pembawa pesan dengan urutan 3"-AAUCCAAAU-5", yang melengkapi satu setengah pengulangan dari enam DNA telomerik (5"-GGGTTA-3"). Fragmen urutan 3'-AAU dari RNA telomerase mengikat ke ujung terminal untai tertinggal template TTA-5', meninggalkan sisa RNA bebas. Deoksiribonukleotida kemudian dilekatkan pada RNA pembawa pesan ini, sehingga memperpanjang urutan berulang dalam DNA dengan satu segmen.
Kemudian telomerase dibelah dan dikirim ke ujung terminal lainnya dengan urutan TTA-5", dan proses diulangi. Segera setelah pengulangan terminal yang cukup lama muncul, DNA polimerase a menempel pada fragmen untai tunggal yang dihasilkan dan melengkapi untai kedua dengan metode komplementaritas dalam arah proksimal 5"-3"-, bergerak menuju situs untai ganda yang sudah ada, fusi berikutnya yang terjadi karena aksi DNA ligase.
Mekanisme perbaikan DNA
Terkadang dalam pertumbuhan rantai misbase secara tidak sengaja terjepit, tetapi untungnya sel yang sehat memiliki enzim perbaikan pasca-replikasi dan sistem koreksi ketidakcocokan dasar yang memperbaiki kesalahan tersebut. Mekanisme kerja sistem ini didasarkan pada penghapusan dan penggantian basa yang dimasukkan secara salah sesuai dengan urutan rantai matriks. Mereka membutuhkan DNA polimerase b dan e untuk berfungsi.
Informasi yang terekam dalam DNA tidak hanya harus diimplementasikan dalam proses perkembangan sel dan organisme, tetapi juga harus sepenuhnya ditransfer ke generasi berikutnya. Untuk itu, sebelum pembelahan sel dilakukan suatu proses di dalamnya. replikasi, yaitu menggandakan jumlah DNA.
Informasi tentang mekanisme replikasi terkandung dalam DNA itu sendiri: beberapa gen mengkodekan enzim yang mensintesis prekursor DNA - nukleotida, yang lain - enzim yang menyediakan koneksi nukleotida yang diaktifkan menjadi satu rantai tunggal. Mekanisme replikasi pertama kali didalilkan oleh J. Watson dan F. Crick, yang mencatat bahwa komplementaritas rantai DNA menunjukkan bahwa molekul ini dapat menggandakan dirinya sendiri. Mereka menyarankan bahwa penggandaan membutuhkan pemutusan ikatan hidrogen dan pemisahan rantai, yang masing-masing memainkan peran template dalam sintesis rantai komplementer. Sebagai hasil dari satu tindakan duplikasi, dua molekul DNA untai ganda terbentuk, yang masing-masing memiliki satu untai induk dan satu untai baru (lihat Gambar.).
Mekanismenya bernama replikasi semi-konservatif. Kemudian, sifat matriks dan prinsip replikasi DNA yang didalilkan dikonfirmasi oleh banyak data eksperimen.
Replikasi DNA dimulai pada titik-titik tertentu pada kromosom - tempat inisiasi replikasi (asal). Proses replikasi dilayani oleh sejumlah besar enzim. Peralatan replikasi DNA bakteri, terutama E. coli, telah dipelajari dengan seksama. Fungsi penguraian molekul DNA pada prokariota dilakukan oleh enzim tertentu. helikase , yang menggunakan energi hidrolisis ATP menjadi ADP untuk bekerja. Mereka sering berfungsi sebagai bagian dari kompleks protein yang menggerakkan garpu dan mereplikasi filamen yang tidak dipilin. Protein spesifik lainnya yang mengikat daerah untai tunggal mencegah untaian DNA bergabung kembali. Bagian-bagian ini, menyimpang ke arah yang berbeda, membentuk struktur karakteristik - garpu replikasi (garpu Kearns). Ini adalah bagian dari molekul DNA di mana sintesis untai baru sedang berlangsung. Protein memainkan peran besar dalam mempromosikan garpu jerapah , termasuk dalam kategori isomerase topologi. Ini hanya ditemukan pada bakteri. Gyrase adalah enzim relaksasi yang, dengan menghasilkan pemutusan untai ganda, menghilangkan positif (di depan garpu) dan mendorong pembentukan superkoil negatif (di belakang garpu) dalam DNA santai.
Setiap untai DNA ibu berfungsi sebagai cetakan untuk sintesis molekul anak. Pada satu untai, sintesis berlangsung terus menerus dalam arah dari ujung 5" ke ujung 3". Rantai ini disebut pemimpin. Rantai kedua dengan arah yang berlawanan, disebut lagging, disintesis dalam bentuk fragmen terpisah, yang kemudian diikat silang oleh ligase menjadi molekul kontinu. Fragmen tersebut dinamai ilmuwan Amerika R. Okazaki, yang pertama kali mendalilkan metode sintesis DNA ini, pecahan Okazaki. Selama sintesis, garpu replikasi bergerak di sepanjang template, dan segmen DNA baru secara berurutan tidak dipilin sampai garpu mencapai titik akhir sintesis (titik terminasi).

Sintesis untai DNA baru membutuhkan fragmen RNA kecil sebagai primer, karena enzim utamanya, DNA polimerase, membutuhkan gugus 3 "OH bebas untuk bekerja. Tiga polimerase DNA yang berbeda dengan fungsi yang sama, ditunjuk sebagai polI, polII dan polIII, telah ditemukan pada prokariota. DNA polimerase I telah dipelajari secara menyeluruh. Ini adalah polipeptida tunggal dengan aktivitas multifungsi (polimerase, 3 "→ 5" exonuclease dan 5 "→ 3" exonuclease).Sintesis primer (primer) dilakukan oleh enzim primase, yang kadang-kadang termasuk dalam kompleks - primosom dari 15-20 protein yang mengaktifkan matriks Primer terdiri dari 10 -60 ribonukleotida Setelah enzim kunci sintesis DNA pada E. coli - polIII - menempelkan deoksiribonukleotida pertama ke benih, dihilangkan menggunakan poli, yang memiliki 3 " → 5" aktivitas eksonuklease, yaitu kemampuan untuk membelah nukleotida terminal dari 3" - ujung rantai. Benih juga disintesis di untai tertinggal di awal setiap fragmen Okazaki. Pembelahannya, serta pemanjangan fragmen yang disintesis oleh polIII, dilakukan oleh polI. Peran polII dalam replikasi DNA E.coli masih belum sepenuhnya jelas.

Selama replikasi DNA eukariotik, proses replikasi diperumit oleh adanya protein dalam kromosom. Untuk mengurai DNA, perlu untuk menghancurkan kompleks DNA dan histon yang sangat padat, dan setelah replikasi, kembali memadatkan molekul anak. Pemutusan DNA menyebabkan supercoiling daerah yang terletak di dekat garpu replikasi. Untuk meredakan ketegangan yang dihasilkan dan gerakan bebas garpu, enzim relaksasi khusus bekerja di sini - topoisomerase. Dua jenis topoisomerase telah diidentifikasi dalam berbagai organisme: tipe I dan II. Mereka mengubah tingkat superkoil dan tipe superkoil dengan menghasilkan pemutusan pada satu (topoisomerase tipe I) atau kedua untai DNA (topoisomerase tipe II) dan menghilangkan risiko terjeratnya DNA.
Replikasi DNA bakteri adalah proses dua arah dengan satu situs inisiasi. Sebaliknya, kromosom eukariotik terdiri dari situs replikasi terpisah - replika dan memiliki banyak situs inisiasi. Replika dapat mereplikasi pada waktu yang berbeda dan pada tingkat yang berbeda. Tingkat replikasi DNA pada sel eukariotik jauh lebih rendah daripada pada sel prokariotik. Pada E. coli, kecepatannya kira-kira 1500 bp. per detik, pada eukariota - 10-100 bp. per detik. DNA sirkular untai ganda dari beberapa virus bereplikasi dalam pola cincin bergulir. Dalam hal ini, satu untai DNA diiris di satu tempat oleh enzim tertentu, dan nukleotida mulai menempel pada ujung OH 3 bebas yang dibentuk dengan bantuan enzim polIII. Molekul sirkular internal berfungsi sebagai matriks. untaian iris dipindahkan dan kemudian digandakan seperti rantai tertinggal E. coli untuk membentuk fragmen yang diikat silang oleh ligase.
Asam nukleat memainkan peran penting dalam memastikan aktivitas vital sel-sel organisme hidup. Perwakilan penting dari kelompok senyawa organik ini adalah DNA, yang membawa semua informasi genetik dan bertanggung jawab atas manifestasi fitur yang diperlukan.
Apa itu replikasi?
Dalam proses pembelahan sel, perlu dilakukan peningkatan jumlah asam nukleat di dalam nukleus agar tidak terjadi kehilangan informasi genetik dalam proses tersebut. Dalam biologi, replikasi adalah penggandaan DNA melalui sintesis untaian baru.
Tujuan utama dari proses ini adalah untuk mentransfer informasi genetik ke sel anak dalam bentuk yang tidak berubah tanpa mutasi.
Enzim dan protein replikasi
Duplikasi molekul DNA dapat dibandingkan dengan setiap proses metabolisme dalam sel yang membutuhkan protein yang sesuai. Karena replikasi merupakan komponen penting dari pembelahan sel dalam biologi, oleh karena itu, banyak peptida tambahan yang terlibat di sini.
- DNA polimerase adalah enzim reduplikasi yang paling penting, yang bertanggung jawab untuk sintesis rantai anak.Dalam sitoplasma sel, dalam proses replikasi, keberadaan trifosfat nukleat adalah wajib, yang membawa semua basa nukleat.
Basa-basa ini adalah monomer asam nukleat, sehingga seluruh rantai molekul dibangun dari mereka. DNA polimerase bertanggung jawab atas proses perakitan dalam urutan yang benar, jika tidak, munculnya semua jenis mutasi tidak dapat dihindari.
- Primase adalah protein yang bertanggung jawab untuk pembentukan primer pada untai cetakan DNA. Primer ini juga disebut primer; ia memiliki monomer awal dari mana sintesis lebih lanjut dari seluruh rantai polinukleotida dimungkinkan untuk enzim DNA polimerase. Fungsi ini dilakukan oleh primer dan enzim yang sesuai.
- Helicase (helicase) membentuk garpu replikasi, yang merupakan divergensi rantai matriks dengan memutus ikatan hidrogen. Ini memudahkan polimerase untuk mendekati molekul dan memulai sintesis.
- topoisomerase. Jika Anda membayangkan molekul DNA sebagai tali yang dipilin, saat polimerase bergerak di sepanjang rantai, tegangan positif akan terbentuk karena lilitan yang kuat. Masalah ini diselesaikan oleh topoisomerase, enzim yang memutus rantai untuk waktu yang singkat dan membuka seluruh molekul. Setelah itu, area yang rusak dijahit kembali, dan DNA tidak mengalami stres.
- Protein ssb menempel seperti kelompok ke untai DNA di garpu replikasi untuk mencegah pembentukan kembali ikatan hidrogen sebelum akhir proses reduplikasi.
- Ligaz. terdiri dari jahitan fragmen Okazaki pada untai tertinggal dari molekul DNA. Ini terjadi dengan memotong primer dan memasukkan monomer asam deoksiribonukleat asli sebagai gantinya.
Dalam biologi, replikasi adalah proses multi-langkah kompleks yang sangat penting dalam pembelahan sel. Oleh karena itu, penggunaan berbagai protein dan enzim diperlukan untuk sintesis yang efisien dan benar.

mekanisme reduplikasi
Ada 3 teori yang menjelaskan proses duplikasi DNA:
- Yang konservatif mengklaim bahwa satu molekul asam nukleat anak memiliki sifat matriks, dan yang kedua sepenuhnya disintesis dari awal.
- Semi-konservatif diusulkan oleh Watson dan Crick dan dikonfirmasi pada tahun 1957 dalam percobaan pada E. Coli. Teori ini menyatakan bahwa kedua molekul DNA anak memiliki satu untai lama dan satu baru disintesis.
- Mekanisme dispersi didasarkan pada teori bahwa molekul anak memiliki bagian yang berselang-seling di sepanjang panjangnya, yang terdiri dari monomer lama dan baru.
Sebuah model semi-konservatif kini telah terbukti secara ilmiah. Apa yang dimaksud dengan replikasi pada tingkat molekuler? Pada awalnya, helikase memutuskan ikatan hidrogen dari molekul DNA, sehingga membuka kedua rantai untuk enzim polimerase. Yang terakhir, setelah pembentukan benih, memulai sintesis rantai baru dalam arah 5'-3'.
Sifat antiparalelisme DNA adalah alasan utama untuk pembentukan untai terkemuka dan tertinggal. Pada untai terdepan, DNA polimerase bergerak terus menerus, dan pada untai tertinggal membentuk fragmen Okazaki, yang di masa depan akan dihubungkan dengan bantuan ligase.

Fitur replikasi
Berapa banyak molekul DNA dalam nukleus setelah replikasi? Proses itu sendiri menyiratkan penggandaan set genetik sel, oleh karena itu, selama periode sintetis mitosis, set diploid memiliki dua kali lebih banyak molekul DNA. Entri seperti itu biasanya ditandai sebagai 2n 4c.
Selain arti biologis dari replikasi, para ilmuwan telah menemukan aplikasi untuk proses di berbagai bidang kedokteran dan sains. Jika dalam biologi replikasi adalah duplikasi DNA, maka di laboratorium, reproduksi molekul asam nukleat digunakan untuk membuat beberapa ribu salinan.
Metode ini disebut reaksi berantai polimerase (PCR). Mekanisme proses ini mirip dengan replikasi in vivo, oleh karena itu, enzim dan sistem buffer yang serupa digunakan untuk jalannya.

kesimpulan
Replikasi sangat penting secara biologis bagi organisme hidup. Transmisi selama pembelahan sel tidak lengkap tanpa duplikasi molekul DNA, sehingga kerja enzim yang terkoordinasi penting pada semua tahap.
Merupakan molekul hereditas, maka untuk mewujudkan kualitas ini, ia harus menyalin dirinya sendiri dengan tepat dan dengan demikian melestarikan semua informasi yang tersedia dalam molekul DNA asli dalam bentuk urutan nukleotida tertentu. Ini dipastikan oleh proses khusus yang mendahului pembelahan sel mana pun dalam tubuh, yang disebut replikasi DNA.
Inti dari replikasi DNA adalah bahwa enzim khusus memutuskan ikatan hidrogen lemah yang menghubungkan nukleotida dari dua rantai. Akibatnya, untaian DNA terputus, dan basa nitrogen bebas “mencuat” dari setiap untai (penampilan yang disebut garpu replikasi). Sebuah enzim khusus, DNA polimerase, mulai bergerak di sepanjang rantai DNA bebas dari 5- ke 3-terminus (untai utama), membantu untuk bergabung dengan nukleotida bebas, terus-menerus disintesis dalam sel, ke ujung 3 "yang baru disintesis Untai DNA Pada untai kedua DNA (lagging strand ) DNA baru terbentuk dalam bentuk segmen-segmen kecil yang terdiri dari 1000-2000 nukleotida (fragmen Okazaki).
Untuk memulai replikasi fragmen DNA untai ini, sintesis fragmen RNA pendek (fitur karakteristik RNA akan dibahas di bawah) sebagai benih diperlukan, yang menggunakan enzim khusus - RNA polimerase (primase). Selanjutnya, primer RNA dikeluarkan, dan DNA dimasukkan ke dalam celah yang terbentuk menggunakan DNA polimerase I. Dengan demikian, setiap untai DNA digunakan sebagai templat atau templat untuk membangun untai komplementer, dan replikasi DNA bersifat semi-konservatif (yaitu satu untai dalam molekul DNA baru adalah "lama", dan yang kedua adalah baru).
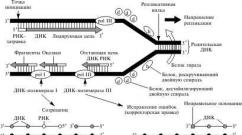
Enzim yang berbeda digunakan untuk mereplikasi untai terkemuka dan tertinggal dalam sel. Sebagai hasil dari replikasi, dua molekul DNA baru yang benar-benar identik terbentuk, yang juga identik dengan molekul DNA asli sebelum dimulainya reduplikasi (proses replikasi DNA ditunjukkan lebih rinci pada Gambar 3.5). DNA polimerase, seperti enzim lainnya, secara signifikan mempercepat proses penambahan nukleotida komplementer ke rantai DNA bebas, namun, afinitas kimia adenin untuk timin, dan sitosin untuk guanin, sangat tinggi sehingga mereka bergabung satu sama lain bahkan tanpa adanya DNA polimerase dalam campuran reaksi sederhana.
Dapat dikatakan, dengan agak disederhanakan, bahwa fenomena penggandaan yang tepat dari molekul DNA, yang didasarkan pada komplementaritas basa-basa molekul ini, adalah basis molekuler dari hereditas. Tingkat replikasi DNA pada manusia relatif lambat, dan akan memakan waktu berminggu-minggu untuk mereplikasi DNA dari setiap kromosom manusia jika replikasi dimulai dari satu titik. Faktanya, dalam molekul DNA dari setiap kromosom, dan setiap kromosom manusia hanya mengandung satu molekul DNA, ada banyak tempat inisiasi replikasi (replika). Dari setiap replika, replikasi berlangsung di kedua arah hingga replika tetangga bergabung. Oleh karena itu, replikasi DNA di setiap kromosom berlangsung relatif cepat.
Replikasi (dari lat. replicatio - pembaruan) - proses sintesis molekul DNA anak pada matriks molekul DNA induk. Selama pembelahan sel induk berikutnya, setiap sel anak menerima satu salinan molekul DNA yang identik dengan DNA sel induk asli. Proses ini memastikan transmisi informasi genetik yang akurat dari generasi ke generasi. Replikasi DNA dilakukan oleh kompleks enzim yang kompleks, terdiri dari 15-20 protein berbeda, yang disebut replisome (bahasa Inggris replisome)
Replikasi DNA adalah peristiwa kunci dalam proses pembelahan sel. Penting bahwa pada saat pembelahan, DNA sepenuhnya direplikasi dan hanya sekali. Ini disediakan oleh mekanisme tertentu dari regulasi replikasi DNA.
Replikasi terjadi di tiga tahap:
1. Inisiasi replikasi
2. Perpanjangan
3. Penghentian replikasi.
Replikasi diatur terutama pada tahap inisiasi. Ini cukup mudah diterapkan, karena replikasi tidak dapat dimulai dari segmen DNA mana pun, tetapi dari segmen yang ditentukan secara ketat, yang disebut tempat inisiasi replikasi. Dalam genom, hanya ada satu atau banyak situs seperti itu. Gagasan situs inisiasi replikasi terkait erat dengan gagasan replika. Replicon adalah hamparan DNA yang berisi situs inisiasi replikasi dan bereplikasi setelah dimulainya sintesis DNA dari situs ini. Genom bakteri, sebagai aturan, adalah replika tunggal, yang berarti bahwa replikasi seluruh genom adalah hasil dari hanya satu tindakan inisiasi replikasi.
Genom eukariotik (serta kromosom individualnya) terdiri dari sejumlah besar replika independen, yang secara signifikan mengurangi total waktu replikasi kromosom individu. Mekanisme molekuler yang mengontrol jumlah inisiasi replikasi di setiap situs per siklus pembelahan sel disebut kontrol jumlah salinan. Selain DNA kromosom, sel bakteri sering mengandung plasmid, yang merupakan replika individu. Plasmid memiliki mekanisme kontrol jumlah salinannya sendiri: mereka dapat menyediakan sintesis hanya satu salinan plasmid per siklus sel, atau ribuan salinan.
Replikasi dimulai di tempat inisiasi replikasi dengan pelepasan heliks ganda DNA, membentuk garpu replikasi, tempat replikasi DNA langsung. Setiap situs dapat membentuk satu atau dua garpu replikasi, tergantung pada apakah replikasi itu searah atau dua arah. Replikasi dua arah lebih umum. Beberapa saat setelah dimulainya replikasi, mikroskop elektron dapat digunakan untuk mengamati mata replikasi - suatu wilayah kromosom di mana DNA telah direplikasi, dikelilingi oleh wilayah DNA yang tidak direplikasi yang lebih luas.
Di garpu replikasi, DNA menyalin kompleks protein besar (replisom), enzim kuncinya adalah DNA polimerase. Garpu replikasi bergerak dengan kecepatan sekitar 100.000 pasangan basa per menit pada prokariota dan 500-5000 pada eukariota.
Mekanisme molekuler replikasi:
Enzim (helicase, topoisomerase) dan protein pengikat DNA melepaskan DNA, menahan matriks dalam keadaan encer, dan memutar molekul DNA. Kebenaran replikasi dipastikan oleh kecocokan yang tepat dari pasangan basa komplementer dan aktivitas DNA polimerase, yang mampu mengenali dan memperbaiki kesalahan. Replikasi pada eukariota dilakukan oleh beberapa polimerase DNA yang berbeda (berbeda dengan replikasi DNA pada prokariota).
DNA polimerase I bekerja pada untai tertinggal untuk menghilangkan primer RNA dan pra-replikasi situs DNA yang dimurnikan. DNA polimerase III adalah enzim replikasi DNA utama yang mensintesis untai DNA terdepan dan fragmen Okazaki selama sintesis untai tertinggal (Fragmen Okazaki adalah fragmen DNA yang relatif pendek yang terbentuk pada untai tertinggal selama replikasi DNA). Selanjutnya, molekul yang disintesis dipelintir sesuai dengan prinsip supercoiling dan pemadatan DNA lebih lanjut. Sintesis adalah energi intensif.
Rantai molekul DNA menyimpang, membentuk garpu replikasi, dan masing-masing menjadi cetakan di mana rantai komplementer baru disintesis. Akibatnya, dua molekul DNA untai ganda baru terbentuk, identik dengan molekul induk.
Dalam proses replikasi, heliks ganda DNA, yang terdiri dari dua rantai polinukleotida komplementer, terurai menjadi rantai terpisah dan, pada saat yang sama, sintesis rantai polinukleotida baru dimulai; dalam hal ini, rantai DNA awal berperan sebagai cetakan. Untai baru yang disintesis pada masing-masing untai asli identik dengan untai asli lainnya. Ketika proses selesai, dua heliks ganda identik terbentuk, yang masing-masing terdiri dari satu rantai lama (asli) dan satu rantai baru (Gbr. 1). Jadi, hanya satu dari dua untai yang membentuk molekul DNA asli yang ditransmisikan dari satu generasi ke generasi lainnya - yang disebut mekanisme replikasi semi-konservatif.
Replikasi terdiri dari sejumlah besar tahap berturut-turut, yang meliputi pengenalan titik awal replikasi, pelepasan dupleks asli (heliks), menjaga agar rantai tetap terisolasi satu sama lain, memulai sintesis rantai anak baru pada mereka, pertumbuhannya (pemanjangan). ), memutar rantai menjadi spiral dan penghentian (akhir) sintesis. Semua tahap replikasi ini, yang berlangsung dengan kecepatan tinggi dan akurasi yang luar biasa, disediakan oleh kompleks yang terdiri dari lebih dari 20 enzim dan protein, yang disebut sistem replikasi DNA, atau replisome. Unit fungsional replikasi adalah replika, yang merupakan segmen (bagian) dari kromosom atau DNA ekstrakromosomal, dibatasi oleh titik awal di mana replikasi dimulai dan titik akhir di mana replikasi berhenti. Tingkat replikasi dikendalikan pada tahap inisiasi. Setelah dimulai, replikasi berlanjut hingga seluruh replicon digandakan (digandakan). Frekuensi inisiasi ditentukan oleh interaksi protein pengatur khusus dengan asal replikasi. Kromosom bakteri mengandung replika tunggal: inisiasi pada satu asal replikasi menyebabkan replikasi seluruh genom. Dalam setiap siklus sel, replikasi dimulai hanya sekali. Plasmid dan virus, yang merupakan elemen genetik otonom, adalah replika terpisah yang mampu melakukan inisiasi ganda dalam sel inang. Kromosom eukariotik (kromosom semua organisme kecuali bakteri dan ganggang biru-hijau) mengandung sejumlah besar replika, yang masing-masing juga dimulai sekali per siklus sel.
Mulai dari titik inisiasi, replikasi terjadi di zona terbatas yang bergerak di sepanjang heliks DNA asli. Zona replikasi aktif ini (yang disebut garpu replikasi) dapat bergerak ke dua arah. Dalam replikasi searah, satu garpu replikasi bergerak di sepanjang DNA. Dalam replikasi dua arah, dua garpu replikasi menyimpang dari titik inisiasi dalam arah yang berlawanan; kecepatan mereka dapat bervariasi. Selama replikasi DNA pada bakteri dan mamalia, laju pertumbuhan rantai anak adalah resp. 500 dan 50 nukleotida dalam 1 detik; pada tumbuhan, nilai ini tidak melebihi 20 nukleotida per detik. Pergerakan dua garpu dalam arah yang berlawanan menciptakan lingkaran yang terlihat seperti "gelembung" atau "mata". Replikasi lanjutan memperluas "mata" sampai mencakup seluruh replika.
Selama replikasi, pertumbuhan rantai dilakukan karena interaksi deoksiribonukleosida trifosfat dengan nukleotida terminal 3 "-OH dari bagian DNA yang sudah dibangun; dalam hal ini, pirofosfat dipecah dan ikatan fosfodiester terbentuk, pertumbuhan rantai polinukleotida hanya terjadi dari ujung 3”nya, yaitu pada arah 5” : 3”. Enzim yang mengkatalisis reaksi ini adalah DNA polimerase.
Energi yang dikeluarkan untuk pembentukan setiap ikatan fosfodiester baru dalam rantai DNA disediakan oleh pemutusan ikatan fosfat antara gugus a- dan b-fosfat dari nukleosida trifosfat.
DNA polimerase memiliki satu situs pengikatan nukleosida trifosfat yang umum untuk keempat nukleotida. Pemilihan nukleotida dari lingkungan, yang basanya komplementer dengan basis cetakan berikutnya, berlangsung tanpa kesalahan karena pengaruh penentuan cetakan DNA (rantai DNA asli). Dengan beberapa kerusakan mutasi pada struktur DNA polimerase, dalam beberapa kasus, masuknya nukleotida non-komplementer terjadi.
Dalam proses replikasi DNA formal, bentuk tautomerik langka dari semua 4 basa nitrogen nukleotida muncul untuk waktu yang singkat dengan probabilitas 10-4-10-5, yang membentuk pasangan tidak beraturan. Akurasi replikasi yang tinggi (probabilitas kesalahan tidak melebihi 10-9) disebabkan oleh adanya mekanisme yang melakukan koreksi (perbaikan).
Garpu replikasi tidak simetris. Dari dua untai anak DNA yang disintesis, satu dibangun terus menerus, dan yang lainnya sebentar-sebentar. Yang pertama disebut yang terdepan, atau yang memimpin, rantai, dan yang kedua disebut yang tertinggal. Sintesis rantai kedua lebih lambat; meskipun, secara keseluruhan, rantai ini dibangun ke arah 3" : 5", masing-masing fragmennya tumbuh secara individual ke arah 5" : 3". Karena mekanisme sintesis yang terputus ini, replikasi kedua rantai antiparalel dilakukan dengan partisipasi satu enzim, DNA polimerase, yang mengkatalisis perpanjangan rantai nukleotida hanya dalam arah 5" : 3".
Sebagai benih untuk sintesis fragmen untai tertinggal, segmen pendek RNA, melengkapi untai cetakan DNA, berfungsi. Primer RNA ini (primer), terdiri dari sekitar 10 nukleotida, disintesis pada interval tertentu pada template untai tertinggal dari ribonukleosida trifosfat dalam arah 5" : 3" menggunakan enzim RNA primase. Primer RNA kemudian diperpanjang dengan deoksinukleotida dari ujung 3' oleh DNA polimerase, yang terus memanjang sampai untai yang dibangun mencapai RNA primer yang melekat pada ujung 5' dari fragmen sebelumnya. Fragmen yang terbentuk dengan cara ini (disebut fragmen Okazaki) dari rantai tertinggal pada bakteri memiliki 1000-2000 residu deoksiribonukleotida; pada sel hewan, panjangnya tidak melebihi 200 nukleotida.
Untuk memastikan pembentukan rantai DNA yang berkesinambungan dari banyak fragmen ini, sistem perbaikan DNA khusus berperan, menghilangkan primer RNA dan menggantinya dengan DNA. Pada bakteri, primer RNA dihilangkan nukleotida oleh nukleotida karena aktivitas eksonuklease DNA polimerase 5':3'. Dalam hal ini, setiap monomer ribonukleotida yang terbelah digantikan oleh deoksiribonukleotida yang sesuai (ujung 3" dari fragmen yang disintesis pada rantai lama digunakan sebagai benih). Enzim DNA ligase melengkapi seluruh proses, mengkatalisis pembentukan fosfodiester ikatan antara gugus 3"-OH dari fragmen DNA baru dan 5" gugus fosfat dari fragmen sebelumnya. Pembentukan ikatan ini membutuhkan energi, yang disuplai selama hidrolisis terkonjugasi ikatan pirofosfat dari koenzim-nikotinamida-adenindinukleotida (dalam sel bakteri) atau ATP (dalam sel hewan dan bakteriofag).
Melepaskan heliks ganda dan spasi. pemisahan rantai dilakukan dengan menggunakan beberapa protein khusus. Helikase membuka bentangan pendek DNA tepat sebelum garpu replikasi. Pemisahan setiap pasangan basa mengkonsumsi energi hidrolisis dua molekul ATP menjadi adenosin difosfat dan fosfat. Beberapa molekul protein pengikat DNA melekat pada masing-masing untai yang terpisah, yang mencegah pembentukan pasangan komplementer dan penyatuan kembali untaian. Karena ini, urutan nukleotida rantai DNA tersedia untuk sistem replikasi. Protein spesifik lainnya membantu primase mengakses templat untai yang tertinggal. Akibatnya, primase mengikat DNA dan mensintesis primer RNA untuk fragmen untai tertinggal. Pembentukan heliks baru tidak memerlukan pengeluaran energi atau partisipasi enzim "pelintir" komplementer.
Dalam kasus replika melingkar (misalnya, dalam plasmid), proses yang dijelaskan disebut q-replikasi. Molekul DNA melingkar dipelintir sendiri (supercoiled), ketika melepas heliks ganda dalam proses replikasi, mereka harus terus berputar di sekitar porosnya sendiri. Dalam hal ini, tegangan puntir muncul, yang dihilangkan dengan memutus salah satu rantai. Kedua ujungnya kemudian segera disambungkan kembali satu sama lain. Fungsi ini dilakukan oleh enzim DNA topoisomerase. Replikasi dalam hal ini biasanya terjadi dalam dua arah, yaitu. ada dua garpu replikasi. Setelah replikasi selesai, dua molekul beruntai ganda muncul, yang pertama-tama dihubungkan satu sama lain sebagai tautan dalam rantai yang sama. Ketika mereka berpisah, salah satu dari dua cincin itu putus untuk sementara.
Replikasi alternatif dari replikon sirkular melibatkan pemutusan salah satu untaian molekul DNA untai ganda. Ujung 3' bebas yang dihasilkan diperpanjang secara kovalen, tetap terikat pada templat (rantai kedua yang tidak terputus), dan ujung 5' secara bertahap digantikan oleh rantai polinukleotida baru. Dengan cara ini, satu rantai terlepas dan terus memanjang sementara garpu replikasi meluncur di sekitar rantai matriks annular (mekanisme cincin bergulir). Saat untai baru tumbuh, untai yang dipindahkan dengan ujung 5" yang dibebaskan menjadi templat linier untuk sintesis untai komplementer baru. Sintesis pada templat linier ini berlanjut hingga untai anak DNA terbentuk yang melengkapi satu putaran dari templat melingkar, yaitu, ke keseluruhan Dengan cara ini, sejumlah besar salinan komplementer dapat diturunkan dari templat melingkar. Mekanisme ini ditemukan di beberapa virus, serta di sejumlah sel eukariotik.
Skema replikasi lain melibatkan pembentukan struktur yang disebut D-loop. Menurut mekanisme ini, pada awalnya hanya satu rantai dari replikon melingkar yang direplikasi, sedangkan rantai kedua, tetap utuh, dipindahkan, membentuk lingkaran. Replikasi untai kedua dimulai dari titik awal yang berbeda dan hanya setelah bagian dari untai pertama telah direplikasi. Mekanisme replikasi seperti itu telah ditemukan, misalnya, dalam DNA mitokondria.
Replikasi RNA (sintesis RNA pada template RNA) kurang dipelajari. Ini dilakukan hanya pada beberapa virus (misalnya, pada virus poliomielitis dan keganasan). Enzim yang mengkatalisis proses ini adalah RNA-dependent RNA polymerase (juga disebut RNA replicase atau RNA synthetase). Ada beberapa jenis replikasi, RNA:
1. virus yang mengandung messenger RNA, atau mRNA [mis. ditelepon (+)RNA], sebagai hasil dari replikasi, mereka membentuk rantai komplementer [(-)RNA], yang bukan mRNA, yang digunakan sebagai cetakan untuk sintesis (+)RNA;
2. virus yang mengandung (--) RNA mensintesis (+) RNA sebagai hasil replikasi;
3. Virus yang mengandung RNA untai ganda [(+)RNA dan (--)RNA] mensintesis (+)RNA sebagai hasil dari replikasi asimetris.
Hipotesis tentang mekanisme replikasi dirumuskan pada tahun 1953 oleh J. Watson dan F. Crick, yang menyarankan bahwa dua untai DNA komplementer setelah pemisahan mereka dapat bertindak sebagai cetakan untuk pembentukan untai DNA baru pada mereka. Pada tahun 1958, M. Meselson dan F. Stahl secara eksperimental mengkonfirmasi mekanisme replikasi ini.