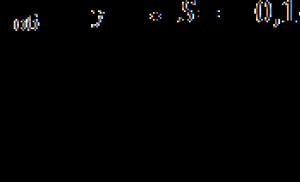Цикл кребса выход атф. Цикл Кребса (цикл трикарбоновых кислот). механизм.Субстраты. Ферменты. Биологические функции цикла Кребса
Цикл Кребса также называется циклом трикарбоновых кислот , так как они образуются в нем в качестве промежуточных продуктов. Представляет собой ферментативный кольцевой конвейер, «работающий» в матриксе митохондрий.
Результатом цикла Кребса является синтез небольшого количества АТФ и образование НАД · H 2 , который далее направляется на следующий этап – дыхательную цепь (окислительное фосфорилирование), расположенную на внутренней мембране митохондрий.
Образовавшаяся в результате пировиноградная кислота (пируват) поступает в митохондрии, где она в конечном итоге полностью окисляется, превращаясь в углекислый газ и воду. Сначала это происходит в цикле Кребса, затем при окислительном фосфорилировании.
До цикла Кребса пируват декарбоксилируется и дегидрируется. В результате декарбоксилирования отщепляется молекула CO 2 , дегидрирование - это отщепление атомов водорода. Они соединяются с НАД.
В результате из пировиноградной кислоты образуется уксусная, которая присоединяется к коферменту А. Получается ацетилкофермент А (ацетил-КоА) – CH 3 CO~S-КоА, содержащий высокоэнергетическую связь.
Превращение пирувата в ацетил-КоА обеспечивает большой ферментативный комплекс, состоящий из десятков полипептидов, связанным с переносчиками электронов.
Цикл Кребса начинается с гидролиза ацетил-КоА, при котором отщепляется ацетильная группа, содержащая два атома углерода. Далее ацетильная группа включается в цикл трикарбоновых кислот.
Ацетильная группа присоединяется к щавелевоуксусной кислоте, имеющей четыре атома углерода. В результате образуется лимонная кислота, включающая шесть атомов углерода. Энергию для этой реакции поставляет макроэргическая связь ацетил-КоА.
Далее следует цепь реакций, в которых связанная в цикле Кребса ацетильная группа дегидрируются с высвобождением четырех пар атомов водорода и декарбоксилируются с образованием двух молекул CO 2 . При этом для окисления используется кислород, отщепляемый от двух молекул воды, а не молекулярный . Процесс называется окислительн ым декарбоксилирование м . В конце цикла щавелевоуксусная кислота регенерируется.
Вернемся на этап лимонной кислоты. Ее окисление проходит за ряд ферментативных реакций, при которых образуются изолимонная, щавелевоянтарная и другие кислоты. В результате этих реакций, на разных стадиях цикла, восстанавливаются три молекулы НАД и одна ФАД, образуется ГТФ (гуанозинтрифосфат), содержащий макроэргическую фосфатную связь, энергия которой впоследствии используется для фосфорилирования АДФ. В результате образуется молекула АТФ.
Лимонная кислота теряет два атома углерода с образованием двух молекул CO 2 .
В результате ферментативных реакций лимонная кислота превращается в щавелевоуксусную, которая снова может соединиться с ацетил-КоА. Цикл повторяется.
В составе лимонной кислоты присоединившийся остаток ацетил-КоА сгорает с образованием углекислого газа, атомов водорода и электронов. Водород и электроны переносятся на НАД и ФАД, которые являются акцепторами для него.
Окисление одной молекулы ацетил-КоА дает одну молекулу АТФ, четыре атома водорода и две молекулы углекислого газа. То есть углекислый газ, выделяемый при аэробном дыхании, образуется на этапе цикла Кребса . При этом молекулярный кислород (O 2) здесь не используется, он необходим лишь на этапе окислительного фосфорилирования.
Атомы водорода присоединяются к НАД или ФАД, в таком виде далее попадают в дыхательную цепь.
Одна молекула глюкозы дает две молекулы пирувата и, следовательно, два ацетил-КоА. Таким образом на одну молекулу глюкозы приходится два оборота цикла трикарбоновых кислот. В общей сложности образуются две молекулы АТФ, четыре CO 2 , восемь атомов H.
Следует отметить, что не только глюкоза и образующийся из нее пируват поступают в цикл Кребса. В результате расщепления ферментом липазой жиров образуются жирные кислоты, окисление которых также приводит к образованию ацетил-КоА, восстановлению НАД, а также ФАД (флавинадениндинуклеотида).
Если клетка испытывает дефицит углеводов и жиров, то окислению могут подвергаться аминокислоты. При этом образуются ацетил-КоА и органические кислоты, которые далее участвуют в цикле Кребса.
Таким образом неважно, каким был первичный источник энергии. В любом случае образуется ацетил-КоА, представляющий собой универсальное для клетки соединение.
Образующийся в ПВК-дегидрогеназной реакции ацетил-SКоА далее вступает в цикл трикарбоновых кислот (ЦТК, цикл лимонной кислоты, цикл Кребса). Кроме пирувата, в цикл вовлекаются кетокислоты, поступающие из катаболизма аминокислот или каких-либо иных веществ.
Цикл трикарбоновых кислот
Цикл протекает в матриксе митохондрий и представляет собой окисление молекулы ацетил-SКоА в восьми последовательных реакциях.
В первой реакции связываются ацетил и оксалоацетат (щавелевоуксусная кислота) с образованием цитрата (лимонной кислоты), далее происходит изомеризация лимонной кислоты до изоцитрата и две реакции дегидрирования с сопутствующим выделением СО 2 и восстановлением НАД.
В пятой реакции образуется ГТФ, это реакция субстратного фосфорилирования . Далее последовательно происходит ФАД-зависимое дегидрирование сукцината (янтарной кислоты), гидратация фумаровой кислоты до малата (яблочная кислота), далее НАД-зависимое дегидрирование с образованием в итоге оксалоацетата .
В итоге после восьми реакций цикла вновь образуется оксалоацетат.
Последние три реакции составляют так называемый биохимический мотив (ФАД-зависимое дегидрирование, гидратация и НАД-зависимое дегидрирование, он используется для введения кетогруппы в структуру сукцината. Этот мотив также присутствует в реакциях β-окисления жирных кислот . В обратной последовательности (восстановление, де гидратация и восстановление) этот мотив наблюдается в реакциях синтеза жирных кислот .
Функции ЦТК
1. Энергетическая
- генерация атомов водорода для работы дыхательной цепи , а именно трех молекул НАДН и одной молекулы ФАДН2 ,
- синтез одной молекулы ГТФ (эквивалентна АТФ).
2. Анаболическая . В ЦТК образуются
- предшественник гема – сукцинил-SКоА ,
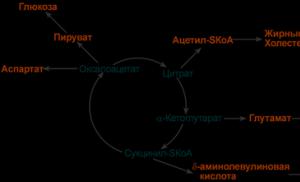
- кетокислоты, способные превращаться в аминокислоты – α-кетоглутарат для глутаминовой кислоты, оксалоацетат для аспарагиновой,
- лимонная кислота , используемая для синтеза жирных кислот ,
- оксалоацетат , используемый для синтеза глюкозы .

Анаболические реакции ЦТК
Регуляция цикла трикарбоновых кислот
Аллостерическая регуляция
Ферменты, катализирующие 1-ю, 3-ю и 4-ю реакции ЦТК, являются чувствительными к аллостерической регуляции метаболитами:
Регуляция доступностью оксалоацетата
Главным и основны регулятором ЦТК является оксалоацетат , а точнее его доступность. Наличие оксалоацетата вовлекает в ЦТК ацетил-SКоА и запускает процесс.
Обычно в клетке имеется баланс между образованием ацетил-SКоА (из глюкозы, жирных кислот или аминокислот) и количеством оксалоацетата. Источником оксалоацетата является пируват , (образуемый из глюкозы или аланина), получение из аспарагиновой кислоты в результате трансаминирования или цикла АМФ-ИМФ, и также поступление из фруктовых кислот самого цикла (янтарной, α-кетоглутаровой, яблочной, лимонной), которые могут образоваться при катаболизме аминокислот или поступать из других процессов.

Синтез оксалоацетата из пирувата
Регуляция активности фермента пируваткарбоксилазы осуществляется при участии ацетил-SКоА . Он является аллостерическим активатором фермента, и без него пируваткарбоксилаза практически неактивна. Когда ацетил-SКоА накапливается, то фермент начинает работать и образуется оксалоацетат, но, естественно, только при наличии пирувата.
Также большинство аминокислот при своем катаболизме способны превращаться в метаболиты ЦТК, которые далее идут в оксалоацетат, чем также поддерживается активность цикла.

Пополнение пула метаболитов ЦТК из аминокислот
Реакции пополнения цикла новыми метаболитами (оксалоацетат, цитрат, α-кетоглутарат и т.п) называются анаплеротическими .
Роль оксалоацетата в метаболизме
Примером существенной роли оксалоацетата служит активация синтеза кетоновых тел и кетоацидоз плазмы крови при недостаточном количестве оксалоацетата в печени . Такое состояние наблюдается при декомпенсации инсулинзависимого сахарного диабета (СД 1 типа) и при голодании. При указанных нарушениях в печени активирован процесс глюконеогенеза , т.е. образования глюкозы из оксалоацетата и других метаболитов, что влечет за собой снижение количества оксалоацетата. Одновременная активация окисления жирных кислот и накопление ацетил-SКоА запускает резервный путь утилизации ацетильной группы – синтез кетоновых тел . В организме при этом развивается закисление крови (кетоацидоз ) с характерной клинической картиной: слабость, головная боль, сонливость, снижение мышечного тонуса, температуры тела и артериального давления.

Изменение скорости реакций ЦТК и причины накопления кетоновых тел при некоторых состояниях
Описанный способ регуляции при участии оксалоацетата является иллюстрацией к красивой формулировке "Жиры сгорают в пламени углеводов ". В ней подразумевается, что "пламень сгорания" глюкозы приводит к появлению пирувата, а пируват превращается не только в ацетил-SКоА, но и в оксалоацетат. Наличие оксалоацетата гарантирует включение ацетильной группы, образуемой из жирных кислот в виде ацетил-SКоА, в первую реакцию ЦТК.
В случае масштабного "сгорания" жирных кислот, которое наблюдается в мышцах при физической работе и в печени при голодании , скорость поступления ацетил-SКоА в реакции ЦТК будет напрямую зависеть от количества оксалоацетата (или окисленной глюкозы).
Если количество оксалоацетата в гепатоците недостаточно (нет глюкозы или она не окисляется до пирувата), то ацетильная группа будет уходить на синтез кетоновых тел . Такое происходит при длительном голодании и сахарном диабете 1 типа .
Образующийся в ПВК-дегидрогеназной реакции ацетил-SКоА далее вступает в цикл трикарбоновых кислот (ЦТК, цикл лимонной кислоты, цикл Кребса). Кроме пирувата, в цикл вовлекаются кетокислоты, поступающие из катаболизма аминокислот или каких-либо иных веществ.
Цикл трикарбоновых кислот
Цикл протекает в матриксе митохондрий и представляет собой окисление молекулы ацетил-SКоА в восьми последовательных реакциях.
В первой реакции связываются ацетил и оксалоацетат (щавелевоуксусная кислота) с образованием цитрата (лимонной кислоты), далее происходит изомеризация лимонной кислоты до изоцитрата и две реакции дегидрирования с сопутствующим выделением СО 2 и восстановлением НАД.
В пятой реакции образуется ГТФ, это реакция субстратного фосфорилирования . Далее последовательно происходит ФАД-зависимое дегидрирование сукцината (янтарной кислоты), гидратация фумаровой кислоты до малата (яблочная кислота), далее НАД-зависимое дегидрирование с образованием в итоге оксалоацетата .
В итоге после восьми реакций цикла вновь образуется оксалоацетат.
Последние три реакции составляют так называемый биохимический мотив (ФАД-зависимое дегидрирование, гидратация и НАД-зависимое дегидрирование, он используется для введения кетогруппы в структуру сукцината. Этот мотив также присутствует в реакциях β-окисления жирных кислот . В обратной последовательности (восстановление, де гидратация и восстановление) этот мотив наблюдается в реакциях синтеза жирных кислот .
Функции ЦТК
1. Энергетическая
- генерация атомов водорода для работы дыхательной цепи , а именно трех молекул НАДН и одной молекулы ФАДН2 ,
- синтез одной молекулы ГТФ (эквивалентна АТФ).
2. Анаболическая . В ЦТК образуются
- предшественник гема – сукцинил-SКоА ,
- кетокислоты, способные превращаться в аминокислоты – α-кетоглутарат для глутаминовой кислоты, оксалоацетат для аспарагиновой,
- лимонная кислота , используемая для синтеза жирных кислот ,
- оксалоацетат , используемый для синтеза глюкозы .
Анаболические реакции ЦТК
Регуляция цикла трикарбоновых кислот
Аллостерическая регуляция
Ферменты, катализирующие 1-ю, 3-ю и 4-ю реакции ЦТК, являются чувствительными к аллостерической регуляции метаболитами:
Регуляция доступностью оксалоацетата
Главным и основны регулятором ЦТК является оксалоацетат , а точнее его доступность. Наличие оксалоацетата вовлекает в ЦТК ацетил-SКоА и запускает процесс.
Обычно в клетке имеется баланс между образованием ацетил-SКоА (из глюкозы, жирных кислот или аминокислот) и количеством оксалоацетата. Источником оксалоацетата является пируват , (образуемый из глюкозы или аланина), получение из аспарагиновой кислоты в результате трансаминирования или цикла АМФ-ИМФ, и также поступление из фруктовых кислот самого цикла (янтарной, α-кетоглутаровой, яблочной, лимонной), которые могут образоваться при катаболизме аминокислот или поступать из других процессов.
Синтез оксалоацетата из пирувата
Регуляция активности фермента пируваткарбоксилазы осуществляется при участии ацетил-SКоА . Он является аллостерическим активатором фермента, и без него пируваткарбоксилаза практически неактивна. Когда ацетил-SКоА накапливается, то фермент начинает работать и образуется оксалоацетат, но, естественно, только при наличии пирувата.
Также большинство аминокислот при своем катаболизме способны превращаться в метаболиты ЦТК, которые далее идут в оксалоацетат, чем также поддерживается активность цикла.
Пополнение пула метаболитов ЦТК из аминокислот
Реакции пополнения цикла новыми метаболитами (оксалоацетат, цитрат, α-кетоглутарат и т.п) называются анаплеротическими .
Роль оксалоацетата в метаболизме
Примером существенной роли оксалоацетата служит активация синтеза кетоновых тел и кетоацидоз плазмы крови при недостаточном количестве оксалоацетата в печени . Такое состояние наблюдается при декомпенсации инсулинзависимого сахарного диабета (СД 1 типа) и при голодании. При указанных нарушениях в печени активирован процесс глюконеогенеза , т.е. образования глюкозы из оксалоацетата и других метаболитов, что влечет за собой снижение количества оксалоацетата. Одновременная активация окисления жирных кислот и накопление ацетил-SКоА запускает резервный путь утилизации ацетильной группы – синтез кетоновых тел . В организме при этом развивается закисление крови (кетоацидоз ) с характерной клинической картиной: слабость, головная боль, сонливость, снижение мышечного тонуса, температуры тела и артериального давления.
Изменение скорости реакций ЦТК и причины накопления кетоновых тел при некоторых состояниях
Описанный способ регуляции при участии оксалоацетата является иллюстрацией к красивой формулировке "Жиры сгорают в пламени углеводов ". В ней подразумевается, что "пламень сгорания" глюкозы приводит к появлению пирувата, а пируват превращается не только в ацетил-SКоА, но и в оксалоацетат. Наличие оксалоацетата гарантирует включение ацетильной группы, образуемой из жирных кислот в виде ацетил-SКоА, в первую реакцию ЦТК.
В случае масштабного "сгорания" жирных кислот, которое наблюдается в мышцах при физической работе и в печени при голодании , скорость поступления ацетил-SКоА в реакции ЦТК будет напрямую зависеть от количества оксалоацетата (или окисленной глюкозы).
Если количество оксалоацетата в гепатоците недостаточно (нет глюкозы или она не окисляется до пирувата), то ацетильная группа будет уходить на синтез кетоновых тел . Такое происходит при длительном голодании и сахарном диабете 1 типа .
4. Цикл трикарбоновых кислот
Вторым компонентом общего пути катаболизма является ЦТК. Этот цикл был открыт в 1937 г. Кребсом и Джонсоном. В 1948 г. Кеннеди и Ленинджер доказали, что ферменты ЦТК локализованы в матриксе митохондрий.
4.1. Химизм цикла трикарбоновых кислот. Свободную уксусную кислоту невозможно окислить путем дегидрирования. Поэтому она в активной форме (ацетил-КоА) предварительно связывается с оксалоацетатом (ЩУК, щавелевоуксусной кислотой), в результате чего образуется цитрат.
1. Ацетил-КоА соединяется с оксалоацетатом в реакции альдольной конденсации, катализируемой цитратсинтазой . Образуется цитрил-КоА. Цитрил-КоА при участии воды гидролизуется до цитрата и НS -КоА.
2. Аконитат-гидратаза (аконитаза ) катализирует превращение цитрата в изоцитрат через стадию цис-аконитовой кислоты. Аконитаза по механизму действия одновременно гидратаза и изомераза.
3. Изоцитратдегидрогеназа катализирует дегидрирование изолимонной кислоты в оксалосукцинат (щавелевоянтарную кислоту), которая затем декарбоксилируется в 2-оксоглутарат (α-кетоглутарат). Коферментом является НАД + (в митохондриях) и НАДФ + (в цитозоле и митохондриях).
4. 2-Оксоглутаратдегидрогеназный комплекс (α-кетоглутаратдегидрогеназный комплекс) катализирует окислительное декарбоксилирование 2-оксоглутарата в сукцинил-КоА. Мультиферментный 2-оксоглутаратдегидрогеназный комплекс похож на пируватдегидрогеназный комплекс и процесс протекает аналогично окислительному декарбоксилированию пирувата.
5. Сукцинилтиокиназа катализирует расщепление сукцинил-КоА на янтарную кислоту и кофермент А. Энергия расщепления сукцинил-КоА накапливается в виде гуанозинтрифосфата (ГТФ). В сопряженной реакции перефосфорилирования АДФ фосфорилируется в АТФ, а освобождающиеся молекулы ГДФ могут вновь фосфорилироваться (субстратное фосфорилирование ). У растений фермент специфичен к АДФ и АТФ.
6. Сукцинатдегидрогеназа катализирует превращение сукцината в фумаровую кислоту. Фермент стереоспецифичен, является интегральным белком, так как вмонтирован во внутреннюю мембрану митохондрий и в качестве простетических групп содержит ФАД и железосерные белки. ФАДН 2 не отделяется от фермента, а два электрона далее передаются на кофермент Q цепи переноса электронов внутренней мембраны митохондрий.
7.Фумарат-гидратаза (фумараза) катализирует превращение фумаровой кислоты в яблочную (малат) с участием воды. Фермент стереоспецифичен, образует только L -малат.
8.Малатдегидрогеназа катализирует окисление яблочной кислоты в оксалоацетат. Кофермент малатдегидрогеназы - НАД + . Далее оксалоацетат вновь конденсируется с ацетил-КоА и цикл повторяется.
4.2. Биологическое значение и регуляция цикла трикарбоновых кислот. Цикл трикарбоновых кислот – компонент общего пути катаболизма, в котором происходит окисление топливных молекул углеводов, жирных кислот и аминокислот. Большинство топливных молекул поступают в ЦТК в виде ацетил-КоА (рис. 1). Все реакции ЦТК протекают согласованно в одном направлении. Суммарная величина D G 0 ¢ = -40 кДж/моль.
В среде врачей давно бытует крылатая фраза «Жиры горят в пламени углеводов». Ее надо понимать как окисление ацетил-КоА, основным источником которого является β-окисление жирных кислот, после конденсации с оксалоацетатом, образуемой, главным образом, из углеводов (при карбоксилировании пирувата). При нарушениях обмена углеводов или голодании создается дефицит оксалоацетата, ведущий к уменьшению окисления ацетил-КоА в ЦТК.

Рис.1. Роль ЦТК в клеточном дыхании. 1 стадия (ЦТК) извлечение из молекулы ацетил-КоА 8 электронов; 2 стадия (цепи переноса электронов) восстановление двух молекул кислорода и формирование протонного градиента (~36 Н +); 3 стадия (АТФ-синтаза) использование энергии протонного градиента для образования АТФ (~9 АТФ) (Berg J .M ., Tymoczko J .L ., Stryer L . Biochemistry . N -Y : W .H .Freeman and Company , 2002).
Основная метаболическая роль ЦТК может быть представлена в виде двух процессов: 1) серия окислительно-восстановительных реакций, в результате которых ацетильная группа окисляется до двух молекул СО 2 ; 2) четырехкратное дегидрирование, ведущее к образованию 3 молекул НАДН+Н + и 1 молекулы ФАДН 2 . Кислород необходим для функционирования ЦТК опосредованно как акцептор электронов в конце цепей переноса электронов и для регенерации НАД + и ФАД.
Основное значение для регуляции ЦТК имеет синтез и гидролиз АТФ.
1. Изоцитратдегидрогеназа аллостерически активируется АДФ путем повышения сродства фермента к субстрату. НАДН ингибирует этот фермент, замещая НАД + . АТФ также ингибирует изоцитратдегидрогеназу. Важно, что превращения метаболитов в ЦТК требуют на нескольких стадиях НАД + и ФАД, количество которых достаточно только в условиях низкого энергетического заряда.
2. Активность 2-оксоглутаратдегидрогеназного (α-кетоглутаратдегидрогеназного) комплекса регулируется аналогично регуляции пируватдегидрогеназного комплекса. Этот комплекс ингибируется сукцинил-КоА и НАДН (конечными продуктами превращений, катализируемых 2-оксоглутаратдегидрогеназным комплексом). Кроме того, 2-оксоглуттаратдегидрогеназный комплекс ингибируется высоким энергетическим зарядом клетки. Итак, скорость превращений в ЦТК уменьшается при достаточной обеспеченности клетки АТФ (рис. 11.2). У ряда бактерий цитратсинтаза аллостерически ингибируется АТФ посредством повышения Км для ацетил-КоА.
Схема регуляции общего пути катаболизма представлена на рисунке 2.

Рис. 2. Регуляция общего пути катаболизма. Основными молекулами, регулирующими функционирование ЦТК являются АТФ и НАДН. Основными пунктами регуляции являются изоцитратдегидрогеназа и 2-оксоглутаратдегидрогеназный комплекс.
4.3. Энергетическая роль общего пути катаболизма
В общем пути катаболизма из 1 молекулы пировиноградной кислоты образуется 3 молекулы СО 2 в следующих реакциях: при окислительном декарбоксилировании пировиноградной кислоты, при декарбоксилировании изолимонной кислоты и при декарбоксилировании 2-оксоглутаровой кислоты. Всего при окислении 1 молекулы пировиноградной кислоты отнимается пять пар атомов водорода, из них одна пара – от сукцината и поступает на ФАД с образованием ФАДН 2 , а четыре пары – на 4 молекулы НАД + с образованием 4 молекул НАДН+Н + при окислительном декарбоксилировании пировиноградной, 2-оксоглутаровой кислот, дегидрировании изоцитрата и малата. В конечном итоге атомы водорода переносятся на кислород с образованием 5 молекул Н 2 О, а выделившаяся энергия аккумулируется в реакциях окислительного фосфорилирования в виде молекул АТФ.
Общий итог:
1. Окислительное декарбоксилирование пирувата ~ 2,5 АТФ.
2. В ЦТК и сопряженных дыхательных цепях ~ 9 АТФ.
3. В реакции субстратного фосфорилирования ЦТК ~ 1 АТФ.
В ЦТК и сопряженных реакциях окислительного фосфорилирования образуется примерно10 АТФ при окислении ацетильной группы одной молекулы ацетил-КоА
Итого в общем пути катаболизма в результате превращений 1 молекулы пировиноградной кислоты выделяется примерно 12,5 молекул АТФ.
Каждому известно, что для нормальной работы организм нуждается в регулярном поступлении целого ряда питательных веществ, которые нужны для здорового метаболизма и, соответственно, баланса процессов выработки и расходования энергии. Процесс выработки энергии, как известно, протекает в митохондриях, которые благодаря этой особенности и получили название энергетических центров клеток. А последовательность химических реакций, которая позволяет получить энергию для работы каждой клеточки тела, называется циклом Кребса.
Цикл Кребса - чудеса, которые происходят в митохондриях
Энергия, получаемая посредством цикла Кребса (также ЦТК - цикл трикарбоновых кислот), идет на нужды отдельных клеток, которые в свою очередь составляют различные ткани и, соответственно, органы и системы нашего организма. Поскольку без энергии организм попросту не может существовать, митохондрии постоянно работают над тем, чтобы бесперебойно поставлять в клетки необходимую им энергию.
Аденозин трифосфат (АТФ) - именно это соединение является универсальным источником энергии, необходимым для протекания всех биохимических процессов в нашем организме.
ЦТК - это центральный метаболический путь, в результате которого завершается окисление метаболитов:
- жирных кислот;
- аминокислот;
- моносахаридов.
В процессе аэробного распада эти биомолекулы расщепляются на меньшие молекулы, которые используются для получения энергии или синтеза новых молекул.
Цикл трикарбоновых кислот состоит из 8 этапов, т.е. реакций:
1. Образование лимонной кислоты:

2. Образование изолимонной кислоты:

3. Дегидрирование и прямое декарбоксилирование изолимонной кислоты.

4. Окислительное декарбоксилирование α-кетоглутаровой кислоты

5. Субстратное фосфорилирование

6. Дегидрирование янтарной кислоты сукцинат-дегидрогеназой

7. Образование яблочной кислоты ферментом фумаразой

8. Образование оксалацетата

Таким образом, после завершения реакций, которые составляют цикл Кребса:
- одна молекула ацетил-КоА (образованная в результате распада глюкозы) окисляется до двух молекул углекислого газа;
- три молекулы NAD восстанавливаются до NADH;
- одна молекула ФАД восстанавливается до ФАДН 2 ;
- образуется одна молекула ГТФ (эквивалент АТФ).
Молекулы НАДН и ФАДН 2 действуют как переносчики электронов и используются для образования АТФ на следующей стадии метаболизма глюкозы - окислительном фосфорилировании.
Функции цикла Кребса:
- катаболическая (окисление ацетильных остатков топливных молекул до конечных продуктов обмена);
- анаболическая (субстраты цикла Кребса - основа для синтеза молекул, в т.ч. аминокислот и глюкозы);
- интегративная (ЦТК - связующее звено между анаболическими и катаболическими реакциями);
- водорододонорная (поставка 3 НАДН.Н + и 1 ФАДН 2 на дыхательную цепь митохондрий);
- энергетическая.
Недостаток элементов, необходимых для нормального протекания цикла Кребса, может привести к серьезным проблемам в организме, связанным с нехваткой энергии.
Благодаря метаболической гибкости организм способен использовать в качестве источника энергии не только глюкозу, но и жиры, расщепление которых также дает молекулы, образующие пировиноградную кислоту (задействуется в цикле Кребса). Таким образом, протекающий надлежащим образом ЦТК обеспечивает получение энергии и строительных блоков для образования новых молекул.