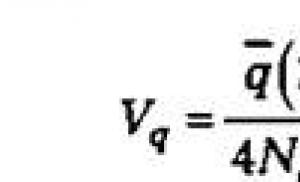Diversitatea genetică a popoarelor. Cum se realizează diversitatea genetică într-o populație Diversitatea genetică reflectă
ÎN POPULAȚIILE UMANE
Omenirea se caracterizează printr-un nivel ridicat de diversitate ereditară, care se manifestă într-o varietate de fenotipuri. Oamenii diferă unul de celălalt prin culoarea pielii, ochilor, părului, forma nasului și a urechii, modelul crestelor epidermice pe vârful degetelor și alte caracteristici complexe. Au fost identificate numeroase variante de proteine individuale, care diferă în unul sau mai multe resturi de aminoacizi și, prin urmare, funcțional. Proteinele sunt trăsături simple și reflectă în mod direct constituția genetică a unui organism. Oamenii nu au aceleași grupe sanguine conform sistemelor de antigen eritrocitar „Rhesus”, AB0, MN. Sunt cunoscute peste 130 de variante ale hemoglobinei și peste 70 de variante ale enzimei glucozo-6-fosfat dehidrogenază (G6PD), care este implicată în descompunerea fără oxigen a glucozei din globulele roșii. În general, cel puțin 30% dintre genele care controlează sinteza enzimelor și a altor proteine la om au mai multe forme alelice. Frecvența de apariție a diferitelor alele ale aceleiași gene variază.
Astfel, dintre numeroasele variante de hemoglobină, doar patru se găsesc în concentrații mari la unele populații: HbS (Africa tropicală, Mediterană), HbS (Africa de Vest), HbD (India), HbE (Asia de Sud-Est). Concentrația altor alele de hemoglobină peste tot aparent nu depășește 0,01-0,0001. Variabilitatea prevalenței alelelor în populațiile umane depinde de acțiunea factorilor evolutivi elementari. Un rol important revine procesului de mutație, selecției naturale, proceselor genetice-automate și migrațiilor.
Procesul de mutație creează noi alele. Și în populațiile umane acționează nedirecționat, aleatoriu. Din această cauză, selecția nu duce la o predominanță pronunțată a concentrației unor alele față de altele. Într-o populație suficient de mare, în care fiecare pereche de părinți din generație în generație produce doi descendenți, probabilitatea de a menține o nouă mutație neutră după 15 generații este de doar 1/9.
Întreaga varietate de variante de proteine, care reflectă diversitatea alelelor din fondul genetic uman, poate fi împărțită în două grupuri. Una dintre ele include variante rare care apar peste tot cu o frecvență mai mică de 1%. Apariția lor se explică numai prin procesul de mutație. Al doilea grup este format din variante întâlnite relativ frecvent în populațiile selectate. Deci, în exemplul cu hemoglobine, primul grup include toate opțiunile cu excepția HbS, HbC, HbD și HbE. Diferențele pe termen lung în concentrația de alele individuale între populații, păstrarea mai multor alele într-o concentrație suficient de mare într-o singură populație, depind de acțiunea selecției naturale sau a derivării genetice.
O formă stabilizatoare a selecției naturale duce la diferențe de interpopulare în concentrația anumitor alele. Distribuția non-aleatorie a alelelor antigenelor eritrocitare AB0 pe planetă se poate datora, de exemplu, ratelor diferite de supraviețuire ale indivizilor care diferă în funcție de grupa sanguină în condiții de epidemii frecvente de infecții deosebit de periculoase. Zonele cu frecvențe relativ scăzute ale alelei I 0 și frecvențe relativ înalte ale alelei I B din Asia coincid aproximativ cu focarele de ciumă. Agentul cauzal al acestei infecții are un antigen asemănător H. Acest lucru face ca persoanele cu grupa sanguină O să fie deosebit de susceptibile la ciuma, deoarece ei, având antigenul H, nu sunt capabili să producă anticorpi anti-ciumă în cantități suficiente. Această explicație este în concordanță cu faptul că concentrații relativ mari ale alelei I 0 se găsesc în populațiile aborigenilor din Australia și Polinezia și indienii americani, care practic nu au fost afectați de ciumă.
Incidența variolei, severitatea simptomelor și mortalitatea sunt mai mari la indivizii cu grupa sanguină A sau AB comparativ cu persoanele cu grupa sanguină 0 sau B (Fig. 12.10). Explicația este că persoanele din primele două grupuri nu au anticorpi care neutralizează parțial antigenul variolic A. Persoanele cu grupa sanguină 0, în medie, sunt capabile să trăiască mai mult, dar au mai multe șanse de a dezvolta ulcer peptic. Tabelul 12.3 identifică unele alele și genotipuri care au semnificație adaptivă în anumite regiuni geografice și de mediu.
Orez. 12.10. Frecvența relativă și indicatori de severitate ai variolei la indivizi cu diferite grupe sanguine ABO
Tabelul 12.3. Exemple de alele cu semnificație adaptivă
| Alele și genotipuri | Distribuție geografică | Sens adaptativ |
| Grupele sanguine ale sistemului AB0, alela B Alela A Transferrine - proteine care leagă fierul, alela Tf DI Fosfataza acidă eritrocitară, alela Acp r alela AcP și grupa sanguină AB alela AcP și grupa sanguină A sau alela AcP și grupa sanguină AB Ceară uscată, alela d | Pretutindeni, mai des în Asia Pretutindeni Frecvență ridicată în zona tropicală Frecvență ridicată printre boșmanii și negroizii din Africa Centrală Frecvență ridicată pe insula Noua Guinee Frecvență ridicată în rândul locuitorilor din Chukotka și Alaska Frecvență ridicată în regiunea Orientului Îndepărtat | Rezistență relativă la ciumă Rezistență relativă la ulcere gastrice și duodenale Rezistență la multe boli infecțioase Activitate enzimatică ridicată la temperaturi ridicate Rezistență la malaria tropicală Ușurință de adaptare în climatele reci Cu genotipul dd, nivelurile de colesterol și concentrațiile de lipide din sânge sunt scăzute, iar lizozimul concentratiile sunt mari in sange.ceara |
În același timp, pentru populațiile din aceeași zonă geografică, dar izolate reproductiv, cauza diferențelor în concentrația alelelor ABO ar putea fi deriva genetică. Astfel, frecvența grupei sanguine A ajunge la 80% în rândul indienilor Blackfoot și la 2% în rândul indienilor din Utah.
Persistența persistentă a mai multor alele ale unei gene în populația umană în același timp se bazează, de regulă, pe selecția în favoarea heterozigoților, ceea ce duce la o stare de polimorfism echilibrat. Un exemplu clasic al acestei situații este distribuția hemoglobinei alelelor S, C și E în focarele de malarie tropicală.
Mai sus sunt exemple de polimorfism la loci specifici, care se explică prin acțiunea unui factor de selecție cunoscut. În condiții naturale, datorită influenței unui complex de factori asupra fenotipurilor organismelor, selecția se realizează în mai multe direcții. Ca urmare, se formează pool-uri de gene care sunt echilibrate în setul și frecvențele alelelor, asigurând supraviețuirea suficientă a populațiilor în aceste condiții. Acest lucru este valabil și pentru populațiile umane. Astfel, persoanele cu grupa sanguină 0 sunt mai susceptibile la ciuma decât persoanele cu grupa B. Tuberculoza pulmonară este tratată cu mai multă dificultate la ei decât la persoanele cu grupa sanguină A. În același timp, tratamentul persoanelor cu sifilis cu grupa sanguină 0 provoacă boala să progreseze mai repede într-un stadiu inactiv. Pentru persoanele cu grupa de sânge 0, probabilitatea de a dezvolta cancer de stomac, cancer de col uterin, reumatism, boală coronariană, colecistită și boli biliare este cu aproximativ 20% mai mică decât pentru persoanele cu grupa A.
Polimorfismul genetic la mulți loci ar putea fi moștenit de oameni de la strămoșii lor în stadiul previzibil de dezvoltare. Polimorfismul în sisteme de grupe de sânge precum AB0 și Rh a fost găsit la maimuțele mari. Factorii de selecție care au creat imaginea actuală a distribuției alelelor în populația umană nu au fost stabiliți cu precizie pentru marea majoritate a loci. Exemplele discutate mai sus indică natura lor ecologică.
Polimorfismul genetic stă la baza variabilității interpopulației și intrapopulației la oameni. Variabilitatea se manifestă în distribuția neuniformă a anumitor boli pe planetă, severitatea apariției lor la diferite populații umane, grade diferite de susceptibilitate a oamenilor la anumite boli, caracteristicile individuale ale dezvoltării proceselor patologice și diferențele de răspuns la efectele terapeutice. . Diversitatea moștenită a fost mult timp un obstacol în calea transfuziei de sânge de succes. În prezent, creează mari dificultăți în rezolvarea problemei transplanturilor de țesuturi și organe.
ÎNCĂRCARE GENETICĂ
ÎN POPULAȚIILE UMANE
Ca și în populațiile altor organisme, diversitatea ereditară reduce capacitatea efectivă a populațiilor umane. Povara genetică a umanității poate fi evaluată prin introducerea conceptului echivalente letale. Se crede că numărul lor per gamet variază de la 1,5 la 2,5 sau de la 3 la 5 per zigot. Aceasta înseamnă că numărul de alele nefavorabile care sunt prezente în genotipul fiecărei persoane, din punct de vedere al efectului lor dăunător total, este echivalent cu efectul a 3-5 alele recesive, ducând în stare homozigotă la moartea individului înainte de vârsta reproductivă.
În prezența alelelor nefavorabile și a combinațiilor lor, aproximativ jumătate din zigoții formați în fiecare generație de oameni sunt incompetenți din punct de vedere biologic. Astfel de zigoți nu participă la transferul genelor către generația următoare. Aproximativ 15% dintre organismele concepute mor înainte de naștere, 3 - la naștere, 2 - imediat după naștere, 3 - mor înainte de a ajunge la pubertate, 20 - nu se căsătoresc, 10% din căsătorii sunt fără copii.
Consecințele adverse ale încărcăturii genetice sub formă de alele recesive, dacă nu duc la moartea organismului, se manifestă printr-o scădere a unui număr de indicatori importanți ai stării individului, în special a abilităților sale mentale. Studiile efectuate asupra populației arabe din Israel, care se caracterizează printr-o frecvență ridicată a căsătoriilor consanguine (34% între verii primari și 4% între verii dubli), au arătat o scădere a abilităților mentale la copiii din astfel de căsătorii.
Perspectivele istorice ale omului, datorită naturii sale sociale, nu sunt legate de informația genetică acumulată de specie Homo sapiens in timpul evolutiei. Cu toate acestea, omenirea continuă să „plătească” pentru aceste perspective, pierzând o parte din membrii săi în fiecare generație din cauza eșecului lor genetic.
Bogăția naturală a planetei noastre provine dintr-o varietate de variații genetice. Diversitatea genetică, adică menținerea heterozigozității genotipice, a polimorfismului și a altor variații genotipice, care este cauzată de nevoia de adaptare a populațiilor naturale, este reprezentată de diversitatea ereditară în interiorul și între populațiile de organisme.
După cum se știe, diversitatea genetică este determinată de variația secvențelor a patru nucleotide complementare din acizii nucleici care alcătuiesc codul genetic. Fiecare specie poartă o cantitate imensă de informații genetice: ADN-ul bacteriilor conține aproximativ 1.000 de gene, ciuperci - până la 10.000, plante superioare - până la 400.000. Există un număr mare de gene în multe plante cu flori și taxoni de animale superioare. De exemplu, ADN-ul unui șoarece de casă conține aproximativ 100.000 de gene.
Noi variații genetice apar la indivizi prin mutații genetice și cromozomiale, precum și în organismele care se caracterizează prin reproducere sexuală, prin recombinare genică. Variațiile genetice pot fi evaluate în oricare
NIVELURI DE BIODIVERSITATE
organisme, de la plante la oameni, ca număr de combinații posibile de diferite forme din fiecare secvență de gene. Alte tipuri de diversitate genetică, cum ar fi cantitatea de ADN per celulă și structura și numărul de cromozomi, pot fi determinate la toate nivelurile de organizare a vieții.
O cantitate imensă de variație genetică este prezentă în populațiile de încrucișare și poate fi determinată prin selecție. Viabilitatea diferită se reflectă în modificările frecvențelor genelor din fondul genetic și este o reflectare reală a evoluției. Semnificația variațiilor genetice este evidentă: ele oferă oportunitatea atât pentru schimbări evolutive, cât și, dacă este necesar, pentru selecția artificială.
Doar o mică parte (aproximativ 1%) din materialul genetic al organismelor superioare a fost studiată într-o măsură suficientă pentru a putea ști care gene sunt responsabile pentru anumite manifestări ale fenotipului organismului. Pentru majoritatea ADN-ului, semnificația acestuia pentru variația formelor de viață rămâne necunoscută.
Fiecare dintre cele 10 9 gene diferite distribuite în întreaga biotă a lumii nu contribuie în mod identic la diversitate. În special, genele care controlează procesele biochimice fundamentale sunt foarte conservate între taxoni și, în general, prezintă o variabilitate scăzută, care este puternic asociată cu viabilitatea organismului.
Dacă pierderea de gene este măsurată din perspectiva ingineriei genetice, având în vedere că fiecare formă de viață este unică, dispariția unei singure specii sălbatice înseamnă pierderea permanentă a mii până la sute de mii de gene cu proprietăți potențiale necunoscute. Ingineria genetică ar putea folosi această diversitate pentru a promova medicina și a crea noi resurse alimentare. Cu toate acestea, distrugerea habitatului și reproducerea limitată a multor specii reduc în mod periculos variația genetică, reducând capacitatea acestora de a se adapta la poluare, schimbări climatice, boli și alte stresuri. Principalul rezervor de resurse genetice - ecosistemele naturale - a fost semnificativ alterat sau distrus.
Scăderea genotipului
Această diversitate care apare sub influența umană pune în pericol posibilitatea unor viitoare adaptări ale ecosistemelor.
Studiul modelelor de distribuție a genotipurilor în populații a fost început de Pearson (1904). El a arătat că în prezența diferitelor alele ale unei gene și a acțiunii încrucișării libere în populații, apare o anumită distribuție a genotipurilor, care poate fi reprezentată ca:
p 2 AA + 2pqAa + p 2 aa,
unde p este concentrația genei A, q este concentrația genei a.
G.H. Hardy (1908) și V. Weinberg (1908), studiind în mod specific această distribuție, și-au exprimat părerea că este echilibru, întrucât în absența factorilor care o perturbă, poate persista în populații timp nelimitat. Așa a început să se dezvolte genetica populației. Principalul merit în dezvoltarea geneticii populațiilor, și mai ales aspectele sale teoretice și matematice, în această perioadă timpurie (1920-1940) îi revine S.S. Chetverikov, S. Wright, R. Fisher, J. Haldane, A.S. Serebrovsky și N.P. Dubinin. *
Evoluția biologică este procesul de acumulare a modificărilor în organism și de creștere a diversității acestora în timp. Schimbările evolutive afectează toate aspectele existenței organismelor vii: morfologia, fiziologia, comportamentul și ecologia acestora. Ele se bazează pe modificări genetice, adică modificări ale substanței ereditare, care, interacționând cu mediul, determină toate caracteristicile organismelor. La nivel genetic, evoluția este acumularea de modificări în structura genetică a populațiilor.
Evoluția la nivel genetic poate fi privită ca un proces în două etape. Pe de o parte, apar mutații și recombinări - procese care determină variabilitatea genetică; pe de altă parte, există deriva genetică și selecția naturală - procese prin care variabilitatea genetică se transmite de la o generație la alta.
Evoluția este posibilă numai dacă există o variație ereditară. Singura sursă de noi variante genetice este procesul de mutație,
NIVELURI DE BIODIVERSITATE
cu toate acestea, aceste variante se pot recombina în moduri noi în timpul reproducerii sexuale, adică în timpul segregării independente a cromozomilor și datorită încrucișării. Variantele genetice care apar ca urmare a proceselor de mutație și recombinare nu se transmit din generație în generație cu succes egal: frecvența unora dintre ele poate crește în detrimentul altora. Pe lângă mutații, procesele care modifică frecvențele alelelor dintr-o populație includ selecția naturală, fluxul de gene (adică migrarea genelor) între populații și deriva genetică aleatorie.
La prima vedere, poate părea că indivizii cu un fenotip dominant ar trebui să fie găsiți mai des decât cu unul recesiv. Cu toate acestea, raportul 3:1 este observat numai la descendenții a doi indivizi heterozigoți pentru aceleași două alele. Cu alte tipuri de încrucișări, la descendenți are loc o împărțire diferită a caracterelor și astfel de încrucișări afectează și frecvențele genotipurilor din populație. Legile lui Mendel nu ne spun nimic despre frecvențele fenotipurilor din populații. Aceste frecvențe sunt discutate în legea Hardy-Weinberg. Principala afirmație a legii Hardy-Weinberg este că, în absența proceselor evolutive elementare, și anume mutația, selecția, migrația și deriva genetică, frecvențele genelor rămân neschimbate de la o generație la alta. Această lege mai spune că, dacă încrucișarea este aleatorie, atunci frecvențele genotipului sunt legate de frecvențele genelor prin relații simple (patratice). Următoarea concluzie rezultă din legea Hardy-Weinberg: dacă frecvențele alelelor la bărbați și femele sunt inițial identice, atunci cu încrucișarea aleatorie frecvențele de echilibru ale genotipurilor la orice locus sunt atinse într-o generație. Dacă frecvențele alelelor celor două sexe sunt inițial diferite, atunci pentru locii autozomali devin aceleași în generația următoare, deoarece atât bărbații, cât și femelele primesc jumătate din genele lor de la tată și jumătate de la mamă. Astfel, frecvențele de echilibru ale genotipurilor sunt realizate în acest caz în două generații. Cu toate acestea, în cazul locurilor legate de sex, frecvențele de echilibru sunt atinse doar treptat.
Legea Hardy-Weinberg a fost formulată în 1908 independent de matematicianul G. H. Hardy din Anglia și de medicul W. Weinberg din Germania. Pentru a înțelege sensul acestei legi, să dăm un exemplu simplu. Să presupunem că acest loc
conține una dintre cele două alele, A și a, prezente la aceleași frecvențe pentru bărbați și femei: p pentru A și q pentru a. Să ne imaginăm că masculii și femelele se încrucișează aleatoriu sau, ceea ce este același lucru, gameții masculilor și femelelor formează zigoți, întâlnindu-se întâmplător. Atunci frecvența oricărui genotip va fi egală cu produsul frecvențelor alelelor corespunzătoare. Probabilitatea ca un anumit individ să aibă genotipul AA este egală cu probabilitatea (p) de a primi alela A de la mamă înmulțită cu probabilitatea (p) de a primi alela A de la tată, adică рхр = р2.
Legea Hardy-Weinberg afirmă că procesul de moștenire nu duce în sine la o schimbare a frecvențelor alelelor și (în cazul încrucișării aleatoare) a frecvențelor genotipului la un anumit locus. Mai mult, cu încrucișarea aleatorie, frecvențele genotipului de echilibru pentru un loc dat sunt atinse într-o generație dacă frecvențele alelelor inițiale sunt aceleași la ambele sexe.
Frecvențele de echilibru ale genotipurilor sunt date de produsele frecvențelor alelelor corespunzătoare. Dacă există doar două alele, A și a, cu frecvențele p și q, atunci frecvențele tuturor celor trei genotipuri posibile sunt exprimate prin ecuația:
(p+q) 2 =p 2 +2pq + q 2 A a AA Aa aa,
unde literele din a doua linie, care desemnează alele și genotipuri, corespund frecvențelor situate deasupra lor în prima linie.
Dacă există trei alele, să spunem A, A2 și A3, cu frecvențele p, q și r, atunci frecvențele genotipului sunt determinate după cum urmează:
(p + q + r) 2 =р 2 + q 2 + r 2 + 2pq+2рг + 2qr А, А г А 3 A, А t A 3 A 2 A 3 A 3 A t A 3, А 2 А 3 A 2 A 3
O tehnică similară de pătrare a unui polinom poate fi utilizată pentru a determina frecvențele de echilibru ale genotipurilor pentru orice număr de alele. Rețineți că suma tuturor frecvențelor alelelor, precum și suma tuturor frecvențelor genotipului, trebuie să fie egală cu unu. Dacă există doar două alele cu frecvențele p și q, atunci p + q - 1 și, prin urmare, p 2 + 2pq + q 2 =(p + q) 2 =1; dacă există trei alele cu oră-
NIVELURI DE BIODIVERSITATE
tots p, q și r, apoi p + q + r = 1 și, prin urmare, de asemenea (p + g + rf = 1 etc.
Organismele cu variante de succes ale trăsăturilor au mai multe șanse decât alte organisme să supraviețuiască și să lase urmași. Ca urmare, variațiile benefice se vor acumula de-a lungul unui număr de generații, iar variațiile dăunătoare sau mai puțin utile vor fi eliminate și eliminate. Acesta se numește procesul de selecție naturală, care joacă un rol principal în determinarea direcției și vitezei evoluției.
Relația directă dintre gradul de variație genetică dintr-o populație și rata de evoluție sub influența selecției naturale a fost dovedită matematic de R. Fisher (1930) în teorema sa fundamentală a selecției naturale. Fisher a introdus conceptul de fitness și a demonstrat că rata de creștere a aptitudinii unei populații în orice moment în timp este egală cu variația genetică a fitnessului în același moment în timp. Cu toate acestea, dovezile directe ale acestui fapt au fost obținute abia la sfârșitul anilor 1960.
Procesul de mutație servește ca sursă a apariției de noi alele mutante și a rearanjamentelor materialului genetic. Cu toate acestea, creșterea frecvenței lor în populație sub influența presiunii mutației are loc extrem de lent, chiar și la scară evolutivă. În plus, marea majoritate a mutațiilor care apar sunt eliminate din populație în câteva generații din motive aleatorii. Inevitabilitatea unui astfel de curs de evenimente a fost fundamentată pentru prima dată de R. Fischer în 1930.
Pentru oameni și alte organisme multicelulare, s-a demonstrat că mutațiile apar de obicei cu o frecvență de 1 la 100.000 (1 10 s) până la 1 la 1.000.000 (1-10 - ®) gameți.
Noi mutanți, deși destul de rari, apar constant în natură, deoarece există mulți indivizi din fiecare specie și mulți loci în genotipul oricărui organism. De exemplu, numărul de indivizi dintr-o anumită specie de insecte este de obicei de aproximativ 100 de milioane (10 8). Dacă presupunem că mutabilitatea medie la un locus este egală cu 1 mutație la 100.000 (10 _ s) gameți, atunci numărul mediu de mutanți noi la acest locus în fiecare generație pentru o anumită specie de insecte va fi de 2-10 8 "10 5 = 2000. ( Frecvența mutațiilor se înmulțește cu numărul de indivizi și cu încă doi, deci
ca orice individ, este produsul fuziunii a doi gameți.) Există aproximativ 100.000 (10 s) de loci în genotipul uman. Să presupunem că oamenii au aceeași rată de mutație ca și Drosophila; în acest caz, probabilitatea ca genotipul fiecărei persoane să conțină o nouă alelă care a fost absentă în genotipul părinților săi este egală cu 2-10 s * 10"® = 2. Cu alte cuvinte, fiecare persoană poartă în medie aproximativ două noi mutații.
Calculele efectuate se bazează pe frecvențele mutațiilor care au manifestări externe. În genomul în ansamblu, rata de mutație este de cel puțin 7-10-9 substituții per pereche de nucleotide pe an. La mamifere, numărul de perechi de nucleotide din genomul diploid este de aproximativ 4*109. În consecință, substituțiile de nucleotide la mamifere apar cu o frecvență de cel puțin 4*108 *7*10“ = 28 pe an per genom diploid. Este clar că procesul de mutație are un potențial enorm de a furniza material ereditar nou.
Un pas important în genetica populației a fost făcut în 1926 de S. S. Chetverikov. Pe baza legii Hardy-Weinberg, Chetverikov a dovedit inevitabilitatea eterogenității genetice în populațiile naturale, având în vedere că noi mutații apar continuu, dar de obicei rămân ascunse (recesive), iar încrucișarea liberă are loc în populație.
Din calculele lui Chetverikov a rezultat și ulterior a fost pe deplin confirmat de practică, că chiar și genele mutante rare și dăunătoare ar fi ascunse în mod sigur de acțiunea purificatoare a selecției naturale la heterozigoți (organisme cu ereditate mixtă) cu gene dominante inofensive de tip sălbatic normal. Mutația va fi, parcă, absorbită de populație, motiv pentru care în spatele uniformității externe a indivizilor unei singure populații, enorma lor eterogenitate genetică va fi inevitabil ascunsă. Chetverikov a exprimat-o astfel: „O specie, ca un burete, absoarbe genovariațiile heterozigote, rămânând tot timpul omogenă din punct de vedere extern (fenotipic). Această caracteristică poate avea două consecințe diferite asupra vieții populațiilor. În marea majoritate a cazurilor, când condițiile de mediu se modifică, o specie își poate realiza „rezerva de mobilizare” a variabilității genetice nu numai datorită noilor modificări ereditare la fiecare individ, ci și datorită „capitalului genetic” moștenit de la strămoșii săi. Datorită acestei blăni-
NIVELURI DE BIODIVERSITATE
Prin moștenire scăzută, o populație dobândește plasticitate, fără de care este imposibil să se asigure stabilitatea adaptărilor în condițiile de mediu în schimbare. Cu toate acestea, ocazional este posibil un alt rezultat: rareori mutații dăunătoare ascunse pot apărea uneori la descendenții părinților complet sănătoși, ducând la apariția unor indivizi cu boli ereditare. Și acesta este și un fenomen biologic firesc, ineradicabil, un fel de plată crudă a populației pentru menținerea eterogenității sale ereditare.
Genetica populației îi datorează lui S.S. Chetverikov o altă descoperire, care a fost conturată într-o mică notă, de doar patru pagini, „Valurile vieții”, publicată în 1905 pe paginile „Jurnalului Departamentului de zoologie al Societății Imperiale a Iubitorilor de Istorie Naturală și Etnografie” din Sankt Petersburg. El a observat că, deoarece orice populație naturală are un număr limitat și limitat de indivizi, acest lucru va duce inevitabil la procese statistice pur aleatorii în răspândirea mutațiilor. În același timp, populațiile tuturor speciilor își schimbă în mod constant dimensiunea (numărul rozătoarelor din pădure se poate schimba de sute de ori de la an la an și de zeci de mii de ori pentru multe specii de insecte), motiv pentru care răspândirea mutațiile în populații pot fi complet diferite în diferiți ani. Dintr-o populație imensă de păsări, insecte, iepuri de câmp și alte animale într-un an dificil, pot rămâne doar câțiva indivizi, uneori complet atipici pentru fosta populație. Dar ei sunt cei care vor da naștere urmași și le vor transmite fondul lor genetic, astfel încât noua populație să fie complet diferită în compoziția materialului genetic față de cea anterioară. Aici se manifestă „efectul fondator” genetic al populației. Genomul populațiilor umane este, de asemenea, în continuă schimbare. K. Ahlström, folosind material din sudul Suediei, a arătat că în populația umană, nu întregul bazin genetic existent este transmis generației următoare, ci doar o parte selectată sau chiar „smulsă” accidental. Astfel, 20% din generația de aici nu a lăsat deloc descendenți, dar 25% dintre părinții care au avut trei sau mai mulți copii au contribuit cu 55% din generația următoare.
Presiunea constantă a mutațiilor și migrarea genelor, precum și separarea genotipurilor mai puțin adaptate biologic la loci polimorfi echilibrați, creează problema așa-numitei încărcături genetice. Conceptul de genetică
A cărui încărcătură a fost introdusă de G. Möller în 1950 în lucrarea „Încărcarea noastră de mutații”. Conform calculelor sale, între 10 și 50% dintre gameții umani conțin cel puțin o mutație nou apărută. Mutațiile slab dăunătoare, dacă apar doar la un heterozigot, pot provoca mai multe daune unei populații decât mutațiile letale complet recesive. Fiecare dintre noi poartă cel puțin opt mutații dăunătoare ascunse în starea heterozigotă. G. Möller, în colaborare cu N. Morton și J. Crowe (1956), a evaluat încărcătura genetică a mutațiilor prin compararea mortalității infantile în eșantioane aleatorii din populații și în familiile în care au avut loc căsătorii între rude. Ei au identificat însăși sarcina mutațională, care apare ca urmare a presiunii mutaționale, și sarcina de segregare ca o consecință a divizării. Ei au propus calcule ale echivalentului letal corespunzător numărului de mutații care împreună dau un rezultat letal. Astfel, unui echivalent letal poate corespunde unei mutații letale, două semi-legale etc. S-a demonstrat că încărcătura genetică medie la om este de 3-5 echivalenți letali.
Yu. P. Altukhov și echipa sa (1989), ca rezultat al unui studiu pe termen lung al stocurilor locale de pește - populații mari izolate unele de altele cu o structură de subpopulație stabilită istoric - au ajuns la concluzia că acestea sunt foarte stabile în timp. si spatiu. Variabilitatea la nivelul subpopulațiilor individuale nu joacă un rol independent și reflectă diferențe locale în acțiunea de selecție datorită eterogenității condițiilor de viață, precum și influenței factorilor aleatoriu. Yu. G. Rychkov și colegii săi au ajuns la o concluzie similară chiar mai devreme când au studiat grupuri izolate de populații umane - populația indigenă din zona circumpolară a Eurasiei. Geneticistul și crescătorul american I.M. Lerner a prezentat ideea homeostaziei genetice încă din 1954, definind-o ca fiind capacitatea unei populații de a-și echilibra structura genetică și de a rezista schimbărilor bruște. Unul dintre mecanismele importante ale homeostaziei genetice este selecția în favoarea heterozigoților, ceea ce duce la un echilibru echilibrat. În același timp, același mecanism provoacă formarea încărcăturii genetice, adică separarea claselor homozigote de indivizi. O astfel de sarcină a fost numită echilibrată
NIVELURI DE BIODIVERSITATE
baie și este considerată ca o plată pentru menținerea heterozigoților clasificați drept elita genetică a populației.
Frecvențele genelor în populații. Au fost create mai multe modele matematice pentru a descrie situații în genetica populației. În 1928, Wahlund a stabilit că, dacă o populație mare este împărțită în K grupe panmictice, atunci într-o astfel de populație se observă un efect similar cu consecințele consangvinizării într-o populație nedivizată: proporția de homozigoți crește cu cantitatea de variație interpopulațională în frecvențele genelor datorită scăderii proporției de heterozigoți.
O contribuție fundamentală la descrierea diferențierii locale a frecvențelor genelor într-o populație subdivizată în termeni de statistici F a fost făcută de S. Wright, care a fundamentat mai mulți coeficienți P ca indicatori ai unei măsuri de diferențiere genetică:
1) F lT - coeficientul de consangvinizare a unui individ raportat la întreaga populație (G);
2) F IS - coeficientul de consangvinizare a unui individ raportat la subpopulația (S);
3) F ST - coeficientul de consangvinizare al subpopulaţiei raportat la întreaga populaţie subdivizată.
Relația dintre aceste mărimi este dată de egalitatea:
Coeficientul F ST a fost propus de S. Wright în 1943 și de atunci a fost utilizat în mod repetat în analiza distribuțiilor frecvenței genelor în populațiile naturale separate. Coeficientul Wright este de mare interes, deoarece ne permite izolarea unor influențe importante ale subdiviziunii populației și structurii genetice. În acest scop, Wright a propus două modele originale de populație: „modelul insulei” și „izolarea prin distanță”.
Model insulă. Există două versiuni cunoscute ale acestui model:
1) subdiviziunea speciei în mai multe subpopulații care se încrucișează liber cu un volum N eficient din punct de vedere genetic, fiecare schimbând gene cu oricare alta cu probabilitate egală și cu aceeași intensitate m;
2) o populație panmictică mare („continentală”), înconjurată de multe colonii mici izolate, diferențiate genetic („insule”), fiecare dintre ele
rykh primește gene de pe „continent” cu o intensitate de t pe generație. Efectele migrației inverse pot fi neglijate.
O măsură a diferențierii aleatorii a subpopulațiilor într-un astfel de sistem este variația intergrup a frecvențelor genelor:

și, prin urmare, condiția de echilibru dintre deriva și migrarea genelor în termeni de statistici P et poate fi scrisă ca

O soluție mai riguroasă cu privire la V q este dată de formula:

Ca o consecință a interacțiunii derivării și migrației, avem o distribuție probabilistică a frecvențelor genelor. În orice moment T reprezintă o funcţie de ![]() ca măsuri ale presiunii migratorii sistematice
ca măsuri ale presiunii migratorii sistematice ![]() - variația selectivă a frecvenței genelor într-o generație datorită izolării, adică deriva aleatorie:
- variația selectivă a frecvenței genelor într-o generație datorită izolării, adică deriva aleatorie:
Dacă notăm cu q t frecvența unei gene din i-a grupă (p, = = q t = 1) și cu q frecvența aceleiași gene într-o populație subdivizată în ansamblu, atunci frecvența medie a genei iar variaţia lui caracteristică acestuia va fi
În consecință, frecvențele zigoților (genotipurilor) sunt egale

NIVELURI DE BIODIVERSITATE
Comparând frecvențele genotipurilor cu frecvențele lor într-o populație caracterizată prin coeficientul de consangvinizare F, obținem relația
Deoarece valoarea F caracterizează populația subdivizată în ansamblu, frecvențele corespunzătoare ale genotipurilor din aceasta sunt egale cu frecvențele care ar fi caracteristice unei populații consangvinizate separate. Cu alte cuvinte, subdiviziunea unei populații în grupuri separate de încrucișare este echivalentă formal cu prezența consangvinizării în întreaga populație.
Formula generală pentru distribuția staționară a frecvențelor genelor în modelul insulei reprezintă funcția p de densitate de probabilitate de următoarea formă:
 і
і
unde p și q sunt frecvențe alelelor în subpopulații; pnq - frecvențele alelelor medii pentru populația subdivizată în ansamblu; N este dimensiunea efectivă a populației; t - coeficientul de migrare;
b) cu efect combinat de izolare, migrare și selecție
unde toate notațiile sunt aceleași ca în expresia anterioară, &W este fitness-ul mediu intralocus al populației
determinată prin însumarea aptitudinii genotipurilor ținând cont de frecvențele acestora.
Distribuțiile staționare pot descrie:
1) distribuția frecvențelor alelelor mai multor loci din aceeași populație în cazul neutralității sau cu aproximativ aceeași presiune de selecție pe fiecare locus;
2) distribuția frecvențelor genelor oricărui locus în generațiile succesive ale aceleiași populații staționare;
3) distribuția frecvențelor alelelor unuia sau mai multor loci într-un set de populații izolate complet sau parțial.
Toate cele trei tipuri sunt echivalente din punct de vedere matematic.
În modelul insular, valoarea coeficientului de migrare a genelor nu depinde de gradul de îndepărtare a populațiilor.S. Wright (1943) și G. Maleko (1955, 1957) au studiat matematic aceeași populație la care intensitatea schimbului. între subpopulații depinde de distanță. Acest model se numește „izolare prin distanță” și presupune o populație distribuită continuu pe o suprafață mare, depășind semnificativ raza activității individuale în perioada reproductivă. Caracteristicile diferențierii locale într-un astfel de sistem depind de mărimea reproductivă sau „vecinația” din care părinții provin aleatoriu, precum și de dimensiunea zonei.
Potrivit lui S. Wright, dimensiunea unui cartier corespunde aproximativ cu numărul de indivizi eficienți genetic din interiorul unui cerc, a cărui rază este egală cu de două ori deviația standard a lungimii migrației într-o direcție într-o anumită generație, adică distanța dintre locurile de naștere ale părinților și urmașii. ‘
Diferențierea este foarte mare când Nn ~ 20, mult mai puțin, dar totuși destul de pronunțată la Nn ~ 200 și aproape corespunde panmixiei când Nn = 2000.
M. Kimura (1953) a propus un alt model de structură a populaţiei. Se numește „modelul scării” și reprezintă o situație intermediară între modelul insulei Wright și modelele populațiilor distribuite continuu de S. Wright și G. Maleko.
Structura de scară a migrației genelor. În acest model, ca și în modelul insulei, se consideră un set de colonii, una
NIVELURI DE BIODIVERSITATE
Cu toate acestea, schimbul de indivizi are loc numai între coloniile învecinate și, astfel, depinde direct de distanța coloniilor unele de altele.
La echilibru, variația interpopulației a frecvențelor genelor

intensitatea migrațiilor între coloniile adiacente, iar m m este presiunea migrației genelor din exterior asupra întregului set de colonii (corespunde coeficientului m din modelul insular al lui S. Wright). Când 0, atunci a = 1 - , P = 0, iar expresia se reduce
la formula lui Wright. Modelul insulei lui Wright este astfel un caz special al modelului ladder în absența schimbului de gene între coloniile învecinate.
O caracteristică critică a subdiviziunii, explorată și teoretic, este capacitatea unor astfel de populații de a susține o diversitate genetică semnificativ mai mare în comparație cu populațiile panmictice de dimensiuni comparabile. Se crede că această diversitate este cea care permite unei populații să răspundă mai eficient la schimbările de mediu și, ulterior, să-și schimbe structura genotipică - o teză care joacă un rol decisiv în conceptul evolutiv al lui Wright, cunoscut sub numele de „teoria echilibrului schimbător”, în care „suprafața” W este reprezentată ca o hartă topografică cu vârfuri și văi pe un singur peisaj de combinații de gene. În acest model, concluzia cea mai importantă este că procesul evolutiv depinde de un echilibru în continuă schimbare între factorii de stabilitate și schimbare și că condiția cea mai favorabilă pentru aceasta este prezența unei structuri fin subdivizate în care se mențin izolarea și comunicarea încrucișată. într-un echilibru adecvat.
Organismele vii din cadrul unei populații variază. Alelele determină diferite trăsături care pot fi transmise de la părinți la urmași. Schimbarea genelor este importantă pentru proces.
Diversitatea genetică care apare într-o populație este aleatorie și nu există un proces de selecție naturală. Selecția naturală rezultă din interacțiunea dintre diversitatea genetică dintr-o populație și mediu.
Mediul determină ce opțiuni sunt mai favorabile. Astfel, trăsăturile mai favorabile sunt transmise urmașilor în viitor.
Cauzele diversității genetice
Diversitatea genetică apare în principal datorită mutației ADN-ului, fluxului de gene (mișcarea genelor de la o populație la alta) și. Deoarece mediul este instabil, populațiile care sunt variabile genetic se vor putea adapta la situații în schimbare mai bine decât cele care nu conțin diversitate genetică.
- Mutația ADN: este o modificare a secvenței ADN. Aceste tipuri de secvențe pot fi uneori benefice organismelor. Majoritatea mutațiilor care duc la modificări genetice provoacă trăsături care nu sunt nici benefice, nici dăunătoare.
- Fluxul genelor: numită și migrație de gene, fluxul de gene introduce noi gene într-o populație pe măsură ce organismele migrează în medii diferite. Noi combinații de gene sunt posibile prin prezența de noi alele în grupul de gene.
- Reproducere sexuală: promovează schimbarea genetică prin producerea diferitelor combinații de gene. este procesul prin care sau sunt create. Variația genetică apare atunci când alelele din gameți se separă și se combină aleatoriu în timpul fertilizării. Recombinarea genetică a genelor are loc și atunci când segmentele de gene se încrucișează sau se rup în timpul meiozei.
Exemple de diversitate genetică
Culoarea pielii unei persoane, culoarea părului, ochii de diferite culori, gropițele și pistruii sunt toate exemple de variații genetice care pot apărea într-o populație. Exemple de modificări genetice la plante includ frunzele modificate și dezvoltarea de flori care seamănă cu insectele pentru a atrage polenizatorii.
Omenirea se caracterizează printr-un nivel ridicat de diversitate ereditară, care se manifestă într-o varietate de fenotipuri. Oamenii diferă unul de celălalt prin culoarea pielii, ochilor, părului, forma nasului și a urechii, modelul crestelor epidermice pe vârful degetelor și alte caracteristici complexe. Au fost identificate numeroase variante de proteine individuale, care diferă în unul sau mai multe resturi de aminoacizi și, prin urmare, funcțional. Proteinele sunt trăsături simple și reflectă în mod direct constituția genetică a unui organism. Oamenii nu au aceleași grupe sanguine conform sistemelor de antigen eritrocitar „Rhesus”, AB0, MN. Sunt cunoscute peste 130 de variante ale hemoglobinei și peste 70 de variante ale enzimei G6PD, care este implicată în descompunerea fără oxigen a glucozei din eritrocite. În general, cel puțin 30% dintre genele care controlează sinteza enzimelor și a altor proteine la om au mai multe forme alelice. Frecvența de apariție a diferitelor alele ale aceleiași gene variază.
Se crede că diversitatea genetică la mulți loci ar fi putut fi moștenită de oameni vii din grupuri ancestrale. Variabilitatea sistemelor grupelor de sânge, cum ar fi AB0 și Rh, a fost găsită la maimuțele mari. Diversitatea moștenită a fost mult timp un obstacol în calea transfuziei de sânge de succes. În prezent, creează mari dificultăți în rezolvarea problemei transplanturilor de țesuturi și organe. Selecția perechilor donor-recipient se realizează prin compararea antigenelor HLA clasa I și II.
Diferențele în prevalența alelelor în populațiile umane moderne au fost, desigur, determinate de acțiunea factorilor evolutivi elementari în timpul evoluției umane. Un rol important revine procesului de mutație, selecției naturale, proceselor genetice-automate și migrațiilor.
Selecția naturală, asigurând adaptabilitatea grupurilor de oameni la diverse condiții de viață, duce și la diferențe de interpopulare, crescând concentrațiile anumitor alele, ceea ce determină polimorfismul genetic al populațiilor.
Agentul cauzal al acestei infecții are un antigen asemănător H. Persoanele cu grupa sanguină O, având același antigen, nu pot produce anticorpi anti-ciumă în cantități suficiente, deci sunt deosebit de sensibile la ciumă. Această explicație este în concordanță cu faptul că concentrații relativ mari ale alelei I0 se găsesc în populațiile aborigenilor din Australia și Polinezia și indienii americani, care practic nu au fost afectați de ciuma. În mod similar, incidența variolei, severitatea simptomelor acestei boli și rata mortalității prin aceasta sunt mai mari la persoanele cu grupa sanguină A sau AB comparativ cu persoanele cu grupa sanguină 0 sau B. Explicația este că persoanele din primele două grupurilor le lipsesc anticorpii, neutralizând parțial antigenul variolic A. Conform expresiei figurative a geneticienilor, epidemiile furioase sunt imprimate în bazinele de gene ale populațiilor umane.
Pe lângă agenții patogeni, evoluția populațiilor umane a fost influențată de alți factori, în special, apariția de noi alimente în alimentație. Se știe că gena care codifică sinteza enzimei lactază, care descompune zahărul din lapte, este activă la toate persoanele aflate în copilărie în perioada hrănirii cu lapte. Cu toate acestea, în timpul procesului de creștere, activitatea acestei gene scade brusc sau se oprește complet. Cu câteva mii de ani în urmă, oamenii au învățat să obțină lapte de la animale domestice și au început să-l folosească pentru hrană tot timpul. Acesta a devenit factorul care a contribuit la consolidarea în fondul genetic al populațiilor care cresc animale și foloseau lapte proaspăt pentru hrană, mutații la una dintre genele reglatoare. Ca urmare a influenței produsului modificat al genei de reglare, gena lactază a început să-și mențină activitatea pe tot parcursul vieții unei persoane. Acum, aproximativ 70% dintre europeni digeră cu ușurință laptele până la vârsta adultă, în timp ce în anumite părți din Africa, Asia Centrală și de Est, doar 30% din populația adultă are o enzimă activă.
Exemplele de mai sus de polimorfism la loci specifici sunt explicate prin acțiunea factorilor de selecție cunoscuți și indică natura lor ecologică. Pentru marea majoritate a locilor nu au fost stabiliți cu precizie factorii de selecție a căror acțiune a creat imaginea actuală a distribuției alelelor în populația umană.
În condiții naturale, datorită influenței unui complex de factori asupra fenotipurilor organismelor, selecția se realizează în mai multe direcții. Rezultatul final a fost determinat de raportul dintre intensitatea diferitelor direcții de selecție. Ca urmare, s-au format pool-uri de gene care au fost echilibrate în setul și frecvențele alelelor, asigurând supraviețuirea suficientă a populațiilor în aceste condiții. În acest caz, adesea acțiunea selecției în direcția care crește stabilitatea unei populații în raport cu un factor a condus la consolidarea în pool-ul său de gene a alelelor care reduc viabilitatea în raport cu un alt factor. De exemplu, o mutație a genei receptorului de vitamina D, care este asociată cu o predispoziție la ospoporoză (o boală caracterizată prin oase fragile), crește rezistența purtătorului său la tuberculoză. Un alt exemplu este mutația genei CFTR, care duce la fibroză chistică, dar protejează organismul de salmoneloză, în special febra tifoidă. Această genă codifică o proteină de pe suprafața celulelor pe care bacteriile Salmonella o folosesc pentru a le pătrunde. La heterozigoți pentru această mutație, fibroza chistică nu se manifestă, iar alela mutantă îi protejează de infecțiile intestinale, făcând dificilă pătrunderea bacteriilor în celule. Astfel, aceeași alelă a unei gene poate fi atât dăunătoare, cât și benefică, în funcție de doza acesteia în genotip, de expunerea purtătorului său la una sau alta influență a mediului etc.
Pe lângă factorii enumerați, migrațiile în masă ale populației și încrucișarea însoțitoare au adus o anumită contribuție la formarea diversității genetice în populațiile umane. Astfel, au fost identificate cinci centre din care a fost efectuat fluxul diferitelor alele genice în pool-urile de gene ale populațiilor europene.
Prima dintre ele se află în Asia Centrală, de unde a avut loc migrația fermierilor către Europa în timpul neoliticului, care a determinat 28% din diversitatea genetică la locuitorii moderni ai acestui continent. Influența așezării din a doua regiune a popoarelor grupului de limbi uralice explică 22% din variația genelor între europeni. 11% din eterogenitatea alelelor este o contribuție a celui de-al treilea centru - interfluviul Volga și Don, de unde nomazii au venit în Europa în 3000 î.Hr. Următoarea contribuție cea mai mare a migrațiilor reflectă probabil răspândirea metropolelor antice grecești în mileniile II și I î.Hr. și este vizibil în mod deosebit în populațiile din Grecia, sudul Italiei și vestul Turciei. Cel de-al cincilea centru al mutațiilor neobișnuite din regiunea vechii țări basci din nordul Spaniei și sudul Franței a contribuit cel mai puțin la diversitatea alelică a europenilor moderni.
Diferențele în diversitatea și frecvența de apariție a alelelor genice în pool-urile de gene ale populațiilor umane stau la baza diferențelor fenotipice interpopulație și intrapopulație între oameni - variabilitate. Variabilitatea se manifestă în distribuția neuniformă a anumitor boli pe planetă, severitatea apariției lor la diferite populații umane, grade diferite de susceptibilitate a oamenilor la anumite boli, caracteristicile individuale ale dezvoltării proceselor patologice și diferențele de răspuns la efectele terapeutice. . Cunoașterea caracteristicilor enumerate pentru umanitate în ansamblu și pentru evaluarea morbidității într-o anumită populație este necesară pentru un medic modern pregătit profesional.
Diferențele de alele ale genei APOE, localizate la om pe cromozomul 19 și implicate în metabolismul colesterolului, afectează semnificativ riscul bolilor cardiovasculare, una dintre principalele cauze de mortalitate. Dintre marea varietate de alele ale acestei gene, trei variante principale sunt cele mai comune: E2, E3, E4. Alela E3 se caracterizează prin cea mai mare activitate funcțională.
Apare la 80% dintre europeni, 39% dintre ei fiind homozigoți. Dacă alelele E4 și E2 sunt prezente în genotipuri, se observă tulburări ale metabolismului lipidic. Pentru alela E4 a fost demonstrată o asociere cu creșterea nivelului de colesterol total și a lipoproteinelor cu densitate mică, iar pentru alela E2, cu creșterea nivelului de trigliceride din serul sanguin. Prezența alelei homozigote E4 la 7% dintre europeni și a alelei E2 la 4% crește semnificativ riscul acestora de a dezvolta boli cardiovasculare. În același timp, un model geografic clar poate fi urmărit în distribuția celor trei variante de gene indicate. De exemplu, la mutarea în nordul Europei, frecvența alelei E4 crește, E3 scade și E2 rămâne constantă. În Suedia și Finlanda, varianta E4 este de 3 ori mai frecventă decât în Italia. Frecvența bolilor discutate crește aproximativ în aceeași proporție. În general, frecvența alelei E4 este semnificativ mai mare în regiunile tropicale și subtropicale decât în zonele cu climă rece. La africani și polinezieni, mai mult de 40% dintre oameni conțin cel puțin o copie a acestei alele, iar în Noua Guinee este mai mult de 50%. Se crede că această distribuție reflectă ponderea alimentelor grase în dieta umană în ultimele milenii. Sănătatea populațiilor care folosesc predominant alimente de origine vegetală nu a depins atât de semnificativ de activitatea acestei gene, ceea ce s-a reflectat în persistența variantei E4 în pool-urile de gene.
Nivelurile de biodiversitate.
Fiecare specie biologică este unică și conține informații despre dezvoltarea unui organism viu (plantă, animal etc.), care are o mare importanță științifică și aplicativă.
Diversitatea genetică este diversitatea genomurilor organismelor vii, adică. variatii genetice. Diversitatea organismelor vii la acest nivel este determinată de proprietățile macromoleculelor biologice, în special ale acizilor nucleici ADN și ARN. Fiecare specie are o anumită cantitate de informații genetice: ADN-ul bacterian alcătuiește 1000 de gene; plante superioare – 400.000; animale - 100 000. Prin urmare, fiecare formă de viață este unică și dispariția unei specii sălbatice înseamnă pierderea a mii până la sute de mii de specii de gene cu proprietăți potențiale necunoscute.
Unul dintre motivele reducerii diversității speciilor este utilizarea irațională a resurselor biologice, care perturbă mecanismele naturale interne de reproducere a animalelor și plantelor în generații succesive care mențin echilibrul cu mediul. Prin urmare, păstrarea diversității genetice a populațiilor de organisme vii, precum și a animalelor de fermă și a soiurilor de plante, este una dintre problemele stringente ale biologiei.
Populațiile de animale și plante constituie obiectul principal al activității economice umane, prin urmare, dacă dorim să folosim rațional resursele biologice ale pământului și ale mării, trebuie să înțelegem procesele care au loc în populații și să anticipăm ce consecințe are intervenția omului în existența acestora. populaţiile pot duce la.
Caracteristicile biologice ale populațiilor de organisme vii sunt derivate ale abilităților lor ereditare, adică. fondurile lor genetice. Baza genetică este informația ereditară totală care este transmisă de la părinți la urmași.
În populațiile de specii de organisme vii care se reproduc sexual, fondul de gene este alcătuit din întreaga diversitate de gene și alele prezente în populație. În orice populație dată, compoziția fondului genetic se poate schimba constant de la o generație la alta. Noile combinații de gene formează genotipuri unice, care duc la diversitatea genetică în populații. Baza diversității genetice este variabilitatea genotipică (heterozigositate, polimorfism etc.), care contribuie la adaptarea organismelor la condițiile de mediu și la supraviețuirea speciilor. Populațiile ale căror gene se modifică continuu de la o generație la alta suferă modificări evolutive și duc la formarea de noi specii și o creștere a biodiversității organismelor vii. Semnificația variației genetice este că ea este baza pentru schimbarea evolutivă.
Un pool de gene static reflectă absența variației genetice între indivizii unei anumite specii și absența schimbărilor evolutive.
Apariția unor noi variații genetice în organismele vii se datorează faptului că acizii nucleici au capacitatea de a schimba în mod spontan structura atunci când sunt expuși la diverși factori de mediu. Acest lucru duce la transformarea genomului organismelor vii - variabilitate ereditară. La acest nivel, diversitatea este creată prin trei procese independente:
Variații genetice care apar spontan - mutații.
Acțiuni ale selecției naturale în populații mixte.
Izolarea geografică și reproductivă.
În natură, la indivizi apar noi variații genetice prin mutații genetice și cromozomiale, iar în organismele care sunt caracterizate prin reproducere sexuală, apar noi variații genetice prin recombinarea genelor.
De asemenea, pot fi create noi variații genetice prin selectarea organismelor vii folosind tehnici de inginerie genetică.
Potrivit oamenilor de știință, până în 2015 biosfera ar putea pierde 10-15% din speciile sale constitutive. Prin urmare, fondul genetic al planetei noastre este supus unei protecții stricte. Pentru a face acest lucru aveți nevoie de:
1. Conservarea diversității genetice a populațiilor existente de organisme vii în procesul de utilizare economică a acestora (management durabil de mediu).
2. Restaurarea acelor populații de organisme vii a căror structură a fost perturbată.
3. Crearea de noi sisteme de populație în acele regiuni în care există condițiile naturale și economice necesare.