Bentos marin (Enciclopedie). Grupuri ecologice de organisme acvatice Ce animale aparțin organismelor bentonice
BENTHOS (din grecescul bentos - adâncime), un ansamblu de organisme care trăiesc pe sol și în solul rezervoarelor marine și continentale. Bentosul este împărțit în plante (fitobentos) și animale (zoobentos). În zoobentos, există animale care trăiesc în sol - infaună (în principal mulți viermi și bivalve poliheți, echiuride, sipunculide, unele echinoderme etc.), care se deplasează de-a lungul suprafeței solului - onfauna (viermi și moluște poliheți, majoritatea echinodermelor, diverse crustacee) , atașate de substrat - epifauna (bureți, hidroizi, anemone de mare și diverse corali, briozoare, ghinde de mare, unele bivalve etc.), precum și care plutesc în apropierea fundului și se scufundă doar periodic în fund - nektobenthos (creveți). , mizide, niște holoturi, pești de fund etc.). După dimensiune, organismele bentos sunt împărțite în macrobentos - de la 5-10 mm și mai mare (marea majoritate a animalelor de jos), meiobentos - de la 0,5 la 5-10 mm (populația stratului superior al solului) și microbentos - mai puțin de 0,5 mm ( bacterii și alte organisme unicelulare). Baza fitobentosului de mică adâncime din mări sunt macrofitele (algele și ierburile marine); Agregările de diatomee bentonice pot juca, de asemenea, un rol semnificativ. În adâncuri, pe lângă animale, trăiesc doar bacterii și ciuperci inferioare. Biomasa bentosului din mări scade odată cu adâncimea: în zonele litorale și sublitorale superioare - până la 5-10 kg/m2 și mai mult, mai adânc în zona sublitorală - sute și zeci de g/m2, în zona batială - grame , în zona abisală - de obicei nu mai mult de 1 g/m2, iar în regiunile centrale ale oceanelor, care sunt sărace în viață - 0,01 g/m2 sau mai puțin. Ponderea apelor de mică adâncime situate în apropierea continentelor (până la 200 m), ocupând mai puțin de 8% din suprafața fundului oceanului, reprezintă aproximativ 60% din biomasa întregului bentos oceanic, iar ponderea apelor abisale (mai adânci decât 3000 m), ocupând 3/4 din suprafața fundului, reprezintă doar mai puțin de 10 %. Biomasa totală de bentos din ocean este estimată la 10-12 miliarde de tone.În unele zone din estul Oceanului Pacific, la o adâncime de 2,5-3 km, au fost descoperite așa-numitele oaze de viață (în 1979) în apropierea punctelor de desfacere ale ape subterane calde (hidroterme). În aceste zone, biomasa bentosului atinge câțiva kg/m2; fauna lor include multe specii de animale necunoscute anterior: bivalve gigantice și reprezentanți ai pogonophora. În corpurile de apă dulce, bentosul este mai sărac din punct de vedere calitativ și cantitativ decât în corpurile de apă de mare. Animale includ protozoare, bureți, viermi rotunzi, viermi oligocheți, lipitori, moluște, crustacee și larvele multor insecte acvatice. Phytobenthos este reprezentat în principal de alge (în special albastru-verde și characeae) și diverse plante cu flori (punga, nuferi, cozi, stuf și multe altele). Benthos servește drept hrană pentru mulți pești, iar în mări, de asemenea, pentru unele pinipede. Multe specii de bentos marin de mică adâncime sunt supuse pescuitului și acvaculturii.
Organisme care trăiesc pe fundul rezervoarelor continentale și al pâraielor; zona de habitat bentonic se numește benthal. Animalele care aparțin bentos se numesc zoobentos, iar plantele sunt numite fitobentos. Bentosul include, de asemenea, mulți protisti (de exemplu, majoritatea foraminiferelor).
Specie Bentos
„Benthos” înseamnă „adâncime” în greacă. Acest nume include organisme care trăiesc pe sol și în solul fundului râurilor, mărilor și oceanelor.
Bentosul este împărțit în animal (zoobenthos) și vegetal (phytobenthos). În zoobentos există animale care trăiesc în pământ și pe pământ, mobile, sedentare și imobile, parțial înfipte în pământ sau atașate de acesta. După metoda lor de hrănire, ei sunt împărțiți în carnivore, ierbivore și cei care se hrănesc cu particule organice.
Printre animalele bentonice se numără mari, mijlocii și mici. Organismele care se mișcă liber de-a lungul fundului includ stelele de mare și crabii. Există organisme care fie plutesc în sus, fie se întind pe fund - lipa, raze. Există și unele foarte sedentare - moluște chiton, scoici, lapea. Stridiile și alte crustacee sunt atașate de fund, iar lanceletele sunt îngropate în sol. Cea mai mare parte a zoobentosului trăiește în zonele de mică adâncime ale mării. Bentosul vegetal este în principal alge. Bentos servește ca hrană pentru mulți pești și alte animale acvatice și este folosit și de oameni (de exemplu, alge, stridii, crabi, unii pești). Exemple de animale bentonice sunt stelele de mare, stridiile, lipa, midii, metiola, mya, castravetele de mare, stele fragile, anemonele și multe altele.
Există, de asemenea, epibentos, organisme care trăiesc pe stratul de suprafață al sedimentelor de fund și endofauna (infauna), organisme care trăiesc direct în interiorul sedimentului de fund. Epibentosul poate fi imobil (sesil) sau în mișcare (vaginal).
Pe baza metodei de obținere a alimentelor, se disting următoarele tipuri de organisme bentonice:
- Mâncătorii de suspensie
- Mâncătorii de pământ
- Raclete
- Filtre
Bentosul este clasificat după mărime în:
- macrobentos, > 1 mm.
- meiobentos,< 1 мм и >32 µm
- microbentos,< 32 мкм
Vezi si
Scrieți o recenzie despre articolul „Benthos”
Legături
- - O selecție de articole pe Oceanographers.ru.
Literatură
- Benthos // Dicționar enciclopedic biologic / capitol. ed. M. S. Gilyarov. - M.: Enciclopedia Sovietică, 1986. - P. 56.
- Benthos / L. A. Zenkevich, T. F. Shchapova // Marea Enciclopedie Sovietică: [în 30 de volume] / cap. ed. A. M. Prohorov. - Ed. a 3-a. - M. : Enciclopedia sovietică, 1969-1978.
Extras care caracterizează Benthos
Când principele Andrei a părăsit palatul, a simțit că tot interesul și fericirea aduse de victorie fuseseră acum abandonate de el și trecute în mâinile indiferente ale ministrului de război și ale adjutantului politicos. Întreaga lui mentalitate s-a schimbat instantaneu: bătălia i s-a părut o amintire veche, îndepărtată.Prințul Andrei a rămas la Brünn cu prietenul său, diplomatul rus Bilibin.
„Ah, dragă prinț, nu există oaspete mai drăguț”, a spus Bilibin, ieșind să-l întâlnească pe prințul Andrei. - Franz, lucrurile prințului sunt în dormitorul meu! - se întoarse spre servitorul care îl desprindea pe Bolkonsky. - Ce, un vestitor al victoriei? Minunat. Și stau bolnav, după cum vezi.
Prințul Andrei, după ce s-a spălat și s-a îmbrăcat, a ieșit în biroul de lux al diplomatului și s-a așezat la cina pregătită. Bilibin se aşeză calm lângă şemineu.
Prințul Andrei, nu numai după călătoria sa, ci și după întreaga campanie, în timpul căreia a fost lipsit de toate conforturile curățeniei și grația vieții, a trăit o plăcută senzație de relaxare printre acele condiții de viață luxoase cu care se obișnuise încă de când copilărie. În plus, după primirea austriecă, i-a făcut plăcere să vorbească, cel puțin nu în rusă (vorbeau franceză), ci cu un rus care, presupunea el, împărtășește dezgustul general rusesc (acum simțit în mod deosebit) față de austrieci.
Bilibin era un bărbat de vreo treizeci și cinci de ani, singur, în aceeași companie cu prințul Andrei. S-au cunoscut la Sankt Petersburg, dar au devenit și mai apropiați la ultima vizită a prințului Andrei la Viena împreună cu Kutuzov. Așa cum prințul Andrei era un tânăr care a promis că va merge departe în domeniul militar, la fel și mai mult a promis Bilibin în domeniul diplomatic. Era încă tânăr, dar nu mai tânăr diplomat, deoarece a început să servească la vârsta de șaisprezece ani, se afla la Paris, la Copenhaga, iar acum ocupa o poziție destul de importantă la Viena. Atât cancelarul, cât și trimisul nostru la Viena l-au cunoscut și l-au prețuit. Nu a fost unul din numărul atât de mare de diplomați cărora li se cere doar merite negative, să nu facă lucruri cunoscute și să vorbească franceza pentru a fi foarte buni diplomați; a fost unul dintre acei diplomați care iubesc și știu să muncească și, în ciuda lenei sale, își petrecea uneori noaptea la birou. A lucrat la fel de bine, indiferent de natura muncii. Nu era interesat de întrebarea „de ce?”, ci de întrebarea „cum?”. Care era problema diplomatică, nu-i păsa; dar să întocmească o circulară, un memoriu sau un raport cu pricepere, exactitate și grație – i-a găsit mare plăcere în acest lucru. Meritele lui Bilibin au fost apreciate, pe lângă lucrările sale scrise, și prin arta sa de a se adresa și de a vorbi în sfere superioare.
Bilibin iubea conversația la fel cum iubea munca, doar atunci când conversația putea fi elegantă. În societate, a așteptat în mod constant ocazia de a spune ceva remarcabil și a intrat în conversație doar în aceste condiții. Conversația lui Bilibin a fost în mod constant presărată de fraze originale, pline de spirit, de interes general.
Bentos (greacă bentos - adâncime) sunt hidrobionți de fund. Ele pot trăi pe suprafața solului și în grosimea acestuia, conform cărora populația de jos este împărțită în epi- și endobentos. Pe baza dimensiunii, se disting organismele micro-, meio- și macrobentos. Microbentosul include organisme cu dimensiuni mai mici de 0,1 mm, meiobentos - de la 0,1 la 2 mm, macrobentos - mai mult de 2 mm. Conform metodei de hrănire, reprezentanții zoobentosului sunt împărțiți în carnivore (carnivore), ierbivore, detritivore (hrănindu-se cu particule organice) etc.
Organismele bentonice sunt în principal animale atașate sau care se mișcă lent (zoobentos: foraminefere, bureți, celenterate, viermi, brahiopode, ascidie, bureți, corali, anelide, crustacee, larve de insecte, echinoderme etc.), mai numeroase în ape puțin adânci. Reprezentanții comuni ai bentosului includ și bacterii, actinomicete, ciuperci și protozoare (în special rizomi și ciliați). În apele puțin adânci, bentosul include și plante (fitobentos: diatomee, alge verzi, brune, roșii, bacterii). La adâncimi unde nu există lumină, fitobentosul este absent. Unele plante cu flori se găsesc de-a lungul coastelor: zostera marina, rupia. Zonele stâncoase ale fundului sunt cele mai bogate în fitobentos. În lacurile cu apă dulce, zoobentosul este mai puțin abundent și divers decât în mare. Este format din protozoare (ciliate, dafnie), lipitori, moluște, larve de insecte etc. Fitobentosul lacurilor este format din diatomee care plutesc liber, alge verzi și albastru-verzi; algele brune și roșii sunt absente. Înrădăcinarea plantelor de coastă în lacuri formează zone clar definite, a căror compoziție și aspect al speciilor sunt în concordanță cu condițiile de mediu din zona de delimitare uscată-apă. În apa corpurilor de apă dulce continentală, în apropierea țărmului, cresc hidrofitele - plante semi-scufundate în apă (vârf de săgeată, aripile albe, stuf, coda, rogoz, trihete, stuf). Ele sunt înlocuite cu hidatofite - plante scufundate în apă, dar cu frunze plutitoare (lotus, linte de rață, capsule de ouă, chilim, takla) și - mai departe - complet scufundate (pondweed, elodea, hara). Hidratofitele includ, de asemenea, plante care plutesc la suprafață (linte de rață).
Grupul ecologic al bentosului include și periphyton (peri - în jur, phyton - plantă) - un set de organisme murdare care se așează pe diverse obiecte și corpurile altor organisme. Compoziția perifitonului include nu numai plante, ci pot fi bureți, briozoare, barnacles (barnacles), bivalve și alte nevertebrate, care duc de obicei un stil de viață sedentar sau sedentar. Organismele Periphyton se așează pe fundul navelor, zgomote, bușteni și alte obiecte plutitoare, pe plante și animale. În unele cazuri, este imposibil să se traseze o graniță clară între bentos și periphyton, de exemplu, în cazul murdării rocilor și a diferitelor obiecte de pe fund.
Adaptarea organismelor acvatice la modul de viață bentonic și periferic se rezumă în primul rând la dezvoltarea mijloacelor de reținere pe un substrat solid, protecția împotriva adormirii prin depunerea sedimentelor în suspensie și dezvoltarea celor mai eficiente metode de mișcare. Este foarte tipic pentru organismele bentonice și perifitone să se adapteze la o tranziție temporară la un stil de viață pelagic în stadiul larvar, care oferă acestor forme sedentare posibilitatea de a se dispersa.
Retenția pe un substrat solid se realizează în diferite moduri. Atașarea la substrat se observă la multe plante, protozoare, bureți, celenterate, viermi, moluște, crustacee și alți hidrobioți. Atașarea poate fi temporară sau permanentă, iar în funcție de mecanismul său - pneumatic (aspirație), sub formă de germinare continuă sau asemănătoare rădăcinii - folosind fire. Atașarea de aspirație este observată, de exemplu, la moluștele Ancylus, lipitori și anemone de mare. Germinarea continuă poate fi calcaroasă (corali), chitinoasă sau asemănătoare cornului (moluște, lipa). Atașarea folosind rădăcini și rizoizi este caracteristică plantelor superioare și multor alge (de exemplu, alge). Atașarea prin fire bisale este caracteristică unui număr de moluște bivalve (midii, midii zebră).
O altă formă de retenție este pătrunderea în substrat: îngroparea parțială sau completă în pământ sau pătrunderea în roca tare prin forare și șlefuire. Multe moluște, echinoderme, viermi, larve de insecte și chiar unii pești sunt capabili să îngroape. De exemplu, niște anghile de mare sapă o groapă pe fundul nisipos, unde se ascund în caz de pericol. Diferiți crabi, creveți, cefalopode și pești (de exemplu, lipa) s-au adaptat, de asemenea, să se îngroape temporar în pământ. Unii bureți, moluște, echinoderme și crustacee pătrund în substraturi solide, distrugându-le mecanic sau chimic (dizolvarea cu acizi).
Ca protecție împotriva acoperirii cu un strat de sedimente, organismele bentonice din diferite grupuri sistematice converg pentru a dezvolta structuri care sunt ridicate deasupra solului datorită formei adecvate a corpului și care se întind în sus în timpul procesului de creștere. Cea mai comună formă de corp a organismelor bentonice atașate este conic, în formă de pâlnie, în formă de ciupercă, în toate cazurile mai subțire dedesubt (bureți, corali solitari, moluște). Crinii de mare au o tulpină lungă cu care se fixează de pământ, iar bureții de sticlă din genul Euplectella arată ca un tub alungit. Odată cu întinderea în sus, protecția împotriva adormirii în suspensie a organismelor atașate este realizată prin așezarea pe substraturi care se ridică deasupra fundului. Cirripede, midii zebră și briozoare cresc pe roci și pietre, pe diverse obiecte și organisme. Plantele sunt salvate de la adormire prin creșterea lor rapidă.
În funcție de gradul de mobilitate, organismele bentonice se împart în vagabonzi (crabi, caracatițe, stele de mare), în mișcare slabă (moluște, arici de mare) și atașate (bureți, briozoare, corali). În general, în acest grup capacitatea de mișcări active este mai puțin pronunțată decât la organismele pelagice. Cu toate acestea, mobilitatea scăzută a speciilor bentonice și perifitone la vârsta adultă este de obicei compensată de mobilitatea ridicată a puieților lor, ducând un stil de viață pelagic. Multe crustacee și larve de insecte migrează în aval de râuri și râuri. Pentru a face acest lucru, ei se ridică în coloana de apă și, după ce înot o anumită distanță, se stabilesc într-un loc nou.
Cele mai semnificative migrații orizontale la vârsta adultă sunt efectuate de crustacee mari. Crabul Kamchatka Paralithodes camtschtica se deplasează toamna la o distanță de până la 200 km de coastă în larg, iar primăvara se întoarce din zonele sale de iernare în apele de coastă. Migrațiile în masă ale homarilor Panularis argus au loc toamna odată cu declanșarea furtunilor cu viteza de 1 km/h și durează câteva zile. La migrare, homarii formează lanțuri de zeci de indivizi, urmând strict unul după altul, atingând cu antenele pe cel din față.
În mări, zoobentosul este reprezentat în principal de foraminifere, bureți, celenterate, nemerteeni, polihete, sipunculide, briozoare, brahiopode, moluște, crustacee, echinoderme, ascidie și pești (Fig. 3.23). Cea mai mare parte a zoobentosului este limitată în zonele de apă puțin adâncă. În zona litorală și în orizontul superior al zonei sublitorale, masa organismelor animale într-o suprafață de 1 m2 poate atinge multe zeci de kilograme (în principal moluște). La adâncimi de până la 100-150 m, biomasa bentosului este de sute și zeci de grame; la o adâncime de 500-1000 m, biomasa bentosului se calculează uneori și în grame, mai adânc - în fracțiuni de gram, la adâncimi mai mari (abisal) - în miligrame. Zonarea verticală se observă și în distribuția bentosului: predomină moluștele și crustaceele în orizonturile superioare, moluștele, polihetele și echinodermele - în orizonturile medii, iar polihetele, crustaceele și echinodermele - în cele mai profunde. Dintre organismele vegetale marine, cea mai mare parte a bentosului din mări este alcătuită din bacterii și alge (diatomee, verde, maro și roșu). Unele plante cu flori sunt, de asemenea, comune în apropierea coastelor: zostera, phyllospadix, ruppia, etc. Cel mai bogat și mai divers fitobentos se află pe zonele stâncoase și stâncoase ale fundului, care servesc drept substrat durabil pentru atașarea algelor.
Orez. 3.23. Diverși reprezentanți ai bentosului marin [Zenkevich, 1976].
Organisme care se deplasează liber de-a lungul fundului: 1 - crab Carcinus\2 - stea de mare Asterias;
3 - pantă; 4 - lipa; 5 - moluște - chiton (Chiton); 6 - scoici (Pecteri); 7 - farfurie (Patella). Organisme atașate sau culcate pe fund: 8 - stridii. Organisme care se înfundă în pământ: 9 - lanceletă; 10-coice Mua. Organisme de perforare a rocii: 11 - Moluște Pholas; 12 - crustacee - ghinda de mare (Balanus); 13 - Faluzie ascidiană; 14- burete de sticla Euplectella; 15 - vierme tub Spirografis; 16 - Tabularia hidroidă; 17- crin de mare
În apropierea coastelor Murmansk, Mării Albe și Orientului Îndepărtat, algele din alge și fucus (maro) produc adesea biomasă de la 15 până la 30 kg pe 1 m2 de fund în zona litorală și orizontul superior al zonei sublitorale. Desișurile de alge roșii Phyllophora sunt cunoscute în partea de nord-vest a Mării Negre la o adâncime de 20-60 m, unde masa sa este în medie de 1,7 kg pe 1 m2 de fund și, în general, de milioane de tone. Pe soluri moi, fitobentosul se dezvolta doar in zonele de apa putin adanci mai mult sau mai putin ferite de actiunea valurilor. Aici este format în principal din plante cu flori (zoster etc.), al căror sistem radicular le permite să prindă rădăcini în soluri nisipoase și mâloase. Distribuția verticală a algelor depinde de compoziția spectrului solar, atingând adâncimi diferite datorită absorbției inegale a razelor de lungimi de undă diferite; În orizontul superior, algele verzi sunt de obicei concentrate, dedesubt - maro, și chiar mai jos - predominant roșii.
O parte semnificativă din bentosul marin este consumată sau utilizată ca materii prime tehnice. Animalele includ moluște (stridii, midii etc.) și crustacee (crabi, creveți, homari etc.). Din cele 1,2 milioane de tone de nevertebrate marine recoltate anual, 62% sunt moluște și 30% sunt crustacee. Multe moluște au valoare alimentară pentru pești și, în plus, produc sidef și perle. Printre alte animale bentonice, bureții și coralii nobili au importanță comercială.
Unele animale de pe fundul mării sunt dăunătoare. În primul rând, aceștia sunt viermi de lemn marini - moluște bivalve teredinide. Multe organisme marine se instalează în număr mare pe fundul navelor, reducând viteza navelor. Plantele marine de bentos sunt folosite ca hrană și materii prime tehnice: varec, ulva, porfir, ahnfeltia, phyllophora și zoster.
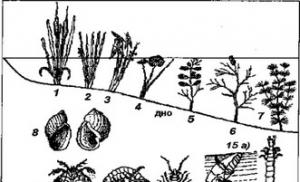
Compoziția bentosului în corpurile de apă dulce continentală este, de asemenea, destul de diversă (Fig. 3.24), dar într-o măsură mai mică decât bentosul marin. Cu toate acestea, lacurile și râurile sunt locuite de numeroase specii de insecte și de larvele acestora, în timp ce în mediul marin speciile de insecte sunt rare. 
Orez. 3.24. Reprezentanți ai bentosului corpurilor de apă dulce continentală.
1 - cattail (Typha); 2 - stuf de lac (Scirpus)\ 3 - vârf de săgeată (Sagittaria);
4- nufar (Nymphaea)\ 5, b-rdest (Potamogeton); 7-alge chara (Chara);
8 - melc de baltă (Ytpaea); 9- acarianul apei (Hydracarina); 10 - crustaceu amfipod (Gammarus); 11 - măgar de apă crustacee izopod (Asellus); 12 - larva de tantari (Culexpipiens); 13 - larva de efei (Cloeon); 14 - larva de zburătoare (Caenis); 15 - gândac de scufundare în apă (Dytiscus); 16-apă scorpion (Ronatra)\ 17-apă smoothie bug (Notonecta);
18 - larva de libelulă (Lestes)
Unele plante care fac parte din bentosul de apă dulce, cum ar fi stuf, stuf etc., sunt folosite în industrie și agricultură. O creștere a cantității de fitobentos de apă dulce se dovedește uneori a fi dăunătoare și duce la creșterea excesivă a piscinei; Mijlocul de control este cosirea. Cu toate acestea, o centură verde de plante scufundate de-a lungul malului unui râu sau al unui lac poate servi ca un filtru biologic eficient - o barieră pentru un corp de apă care poate consuma excesul de substanțe organice și poate împiedica pătrunderea acestora în coloana de apă.
Multe organisme acvatice s-au adaptat existenței la interfața dintre apă și sol. Reprezentanții pelagobenthosului trăiesc în zona de contact a coloanei de apă cu fundul, fie înotând, mișcându-se de-a lungul pământului, fie îngropând în el. În funcție de mărimea și gradul de mobilitate, reprezentanții pelagobenthosului aparțin nekto- sau planktobenthos. Printre reprezentanții nektobenthosului, cei mai des întâlniți sunt racii mai mari și peștii. Planktobentonții tipici includ larve transparente ale țânțarului Chaoborus care trăiesc alternativ în apă și sol, mulți gândaci, copepode și cladocere, o serie de rotifere echipate cu un picior, numeroase alge clorococice, desmidiene și albastre-verzi.
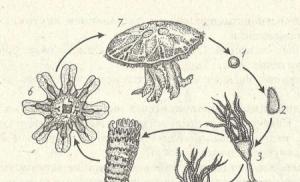
Trebuie remarcat faptul că deseori atribuirea rigidă a organismelor acvatice la unul sau la altul grup ecologic în funcție de habitatul lor este condiționată. Aceeași specie în diferite stadii de ontogeneză pot fi incluse în diferite grupuri ecologice. De exemplu, animalele bentonice tipice, cum ar fi crabii, stelele de mare și moluștele au larve care duc un stil de viață planctonic de ceva timp (Fig. 3.24).
În general, formele de viață ale organismelor acvatice pot fi foarte diverse. În acest caz, ar trebui să se țină cont întotdeauna de lărgimea spectrelor de adaptare ale organismelor. Există abordări de clasificare a locuitorilor acvatici pe baza metodelor lor de hrănire, a relației cu salinitatea, presiunea și temperatura. De asemenea, este posibil să se clasifice animalele acvatice în funcție de modurile lor de mișcare. De exemplu, grupul ecologic de specii care se mișcă în apă într-o manieră reactivă include astfel de reprezentanți eterogene în poziție sistematică precum meduze, cefalopode, unii ciliați și flagelați, larvele unor libelule etc. (Fig. 3.25). 
Orez. 3.25. Reprezentanți ai unui grup ecologic de organisme care se mișcă în apă într-o manieră reactivă.
1 - meduză Cytaeis vulgarus; 2 - Pelagothuria holoturie pelagică;
3 - larva libelulei rocker; 4 - caracatiță înotătoare Octopus vulgaris
Apa este habitatul multor organisme vii. Multe organisme încă trăiesc în ocean sau în corpurile de apă dulce. Oceanul a fost și rămâne o arenă pentru dezvoltarea unei vieți vibrante și diverse. Este locuit de 300 de mii de specii de animale și microorganisme și 30 de mii de specii de plante.
De ce există o diferență atât de mare în compoziția speciilor a florei și faunei din Oceanul Mondial? Faptul este că animalele locuiesc pe toată grosimea apelor oceanului, iar plantele pot trăi doar până la o adâncime de 100 m, unde ajung razele soarelui. Și deși în comparație cu grosimea oceanului, o adâncime de o sută de metri nu pare foarte semnificativă, acest strat este cel care asigură viața în el.
Organismele vii din ocean și corpurile de apă dulce sunt împărțite în trei grupuri: plancton, nectonȘi bentos.
Plancton(din greacă planktos - rătăcitor) sunt toate organismele care locuiesc în corpurile de apă dulce și sărată și sunt aproape incapabile de mișcare independentă (Fig. 167). Ei trăiesc în stare suspendată, ca și cum ar pluti, deoarece nu au organe de mișcare capabile să reziste mișcării apei, cum ar fi curenții. Dimensiunea acestor locuitori variază de la microscopică la câțiva centimetri. Planctonul este împărțit în animale - zooplancton si legume - fitoplancton.
Este clar că reprezentanții fitoplancton-on— algele capabile de fotosinteză locuiesc în oceane și mări doar la o adâncime de 100 m. Fitoplanctonul servește drept hrană pentru zooplancton și alte animale acvatice.
 |
| Orez. 168. Balenă cu cocoașă |
Zooplancton distribuite pe toată coloana de apă. Este reprezentat de mici crustacee, protozoare, celenterate, ouă și larve de pești. Majoritatea organismelor planctonice se caracterizează prin adaptări pentru plutirea în coloana de apă: excrescențe lungi, incluziuni de gaze sau grăsimi, corp gelatinos etc.
Diferiți pești mici se hrănesc cu plancton ( sardine, hamsii si etc.). Și nu numai. Aceasta este o delicatesă preferată a celor mai mari animale marine - balenele(Fig. 168), care absorb cantități enorme din acești cei mai mici locuitori ai oceanului, trecând constant apa prin plăcile de fani.
Nekton(din greacă nektos - plutitor) - animale acvatice care înoată activ, care sunt capabile să reziste forței curentului și să se deplaseze pe distanțe lungi. Acestea includ peşte, calmar, cetacee, pinipede, șerpi de apă, țestoase, pinguini si etc. Material de pe site
Bentos(din greacă bentos - adâncime) - organisme care trăiesc pe sol și în solul fundului râurilor, mărilor și oceanelor. Animalele bentonice care se deplasează activ includ stele de mare, crabi, rac de râu(Fig. 169). Există organisme care fie plutesc, fie zac în fund - cambulă(Fig. 170) și pante. Moluște sedentare ( scoici, farfurioare). Stridiile și alte crustacee sunt atașate de fund și îngropate în sol. lancelete. Cea mai mare parte a bentosului trăiește în zone puțin adânci ale mării.
Pe această pagină există material pe următoarele subiecte:
Informații despre bentos în ocean
Raport despre bentos
Plancton nekton bentos cum se mișcă
Raport asupra plancton necton bentos
Prezentare pe bentos
Întrebări despre acest material:
Benthos (din grecescul bentos - adâncime) - un set de animale și plante care trăiesc pe fund sau asociate cu fundul; Multe dintre aceste organisme trec printr-un stadiu planctonic de dezvoltare, ceea ce facilitează dispersarea lor. Pe baza dimensiunilor predominante ale organismelor care alcătuiesc bentosul, acesta este împărțit în microbentos (bacterii, protozoare, diatomee de fund etc.), meiobentos (viermi mici, crustacee și alte organisme cu o lungime a corpului, de obicei, nu mai mare de 2 mm) și macrobentos (organisme de fund mai mari de 2 mm). Există fitobentos - alge și ierburi marine care locuiesc doar în părțile iluminate ale raftului și zoobentos - animale de fund care locuiesc pe fundul oceanului până la adâncimi ultra-abisale. Organismele zoobentos pot trăi pe talii vegetali, pe suprafața solului (epifauna), pot săpa în pământ relativ moale (infauna) sau să sape în roci fragile (endolitofauna); ele diferă prin gradul de mobilitate (de la forme atașate până la târăre activ). Speciile asociate cu solul, dar capabile să înoate activ, sunt clasificate drept nektobenthos (raya, lipa etc.). Dintre animalele Benthos, unele se hrănesc direct cu plante (fitofage), altele consumă substanțe organice suspendate în stratul inferior al apei (setonofage) sau conținute în sol (detritivore), prădătorii se hrănesc cu animale; în plus, există mâncători de carii (necrofagi) și omnivore. Organismele bentos joacă un rol important în ecosistemele naturale, formând lanțuri alimentare complexe (trofice) și furnizează hrană pentru pești, mamifere și păsări. Mulți reprezentanți ai Benthosului au fost folosiți de mult timp ca hrană de oameni și servesc ca obiecte de pescuit și cultivare.
Fitoplanctonul este baza nutriției pentru zooplancton și bentos. În același timp, dezvoltarea organismelor acvatice depinde de compoziția speciei și de abundența plantelor scufundate și a plantelor cu frunze plutitoare. Cu cât este mai diversă compoziția și structura creșterii excesive a vegetației acvatice, cu atât zooplanctonul este mai divers și mai bogat (Baklanovskaya, 1956).
Se observă o relație inversă între vegetație și biomasa zoobentosului. Productivitatea ridicată a bentosului în corpurile de apă dulce se observă pe soluri slab supraîncărcate, cu o cantitate mică de reziduuri de plante și o acumulare slabă (Vonokov, 1956; Kosova, 1958). Deteriorarea regimului de oxigen într-un rezervor, scăderea afluxului de nutrienți și predominarea proceselor de acumulare a substanțelor organice în balanță duc la dominarea grupelor de plante monospecifice sau epuizate și, în consecință, la scăderea productivității fitoplanctonului. , zooplancton și zoobentos.
Conform observațiilor lui M.V. Pavlova, biomasa medie anuală a bentosului din cele mai productive zone ale lacului Issyk-Kul - golfuri este de 104,2 kg/ha. Pentru comparație: biomasa medie de bentos în zona de coastă a lacului Baikal este de 220 kg/ha. În ceea ce privește productivitatea bentosului, Issyk-Kul este clasificat ca un rezervor cu capacitate medie de hrănire.
- 2. Esența fezabilității biologice.
În interconectarea tuturor fenomenelor Universului se află originile oportunității care domnește în natura vie. Adică, originile fenomenului vieții ar trebui căutate în structura și trăsăturile dezvoltării Universului.
Fenomenul de oportunitate (teleologicitate) este inițial inerent naturii. Se bazează pe principiul optimității, motiv pentru care este unitatea Universului și complementaritatea tuturor proceselor care au loc în el. Iritabilitate, instinct, psihic, rațiune - toate acestea sunt doar câteva dintre cele mai cunoscute manifestări ale fenomenului oportunității pentru noi. Se poate demonstra că toate sunt mecanisme specifice găsite de natură pentru a implementa principiul optimității. Așa că o persoană își folosește mintea pentru a evalua consecințele pașilor lui pentru a găsi cel mai optim comportament. Psihicul animalelor servește și el acestui scop, dar „gradul de pătrundere în viitor” al psihicului este mult mai mic decât cel al minții. Ceea ce înțelegem ca legi specifice ale naturii îndeplinește funcții similare, dar cu un „grad de pătrundere în viitor” și mai nesemnificativ. Astfel, se știe din teoria cuantică că și în procesul de interacțiune a două particule elementare există o fază paradoxală când particulele primesc cumva informații (predicție) despre viitorul apropiat. Principiul optimității se poate baza pe două postulate complementare:
1) orice sistem se străduiește să ocupe o stare în care orice modificare în cadrul sistemului nu are practic niciun efect (afectează modul minim posibil) asupra stării sistemului în ansamblu;
2) dintre toate stările posibile, în fiecare moment în timp, se realizează starea cu care este asociat cel mai mic număr de modificări.
Primul postulat stă la baza dinamicii Universului, făcându-l să evolueze de la stări de neechilibru la stări din ce în ce mai mult echilibrate. Al doilea postulat interzice tranzițiile bruște la stările de echilibru, forțând întotdeauna să construiască lanțuri clare de evenimente cauză-efect. Se poate observa că al doilea postulat apare ca urmare a acțiunii principiului Le Chatelier-Brown (natura încearcă să încetinească orice modificare) atunci când sistemul încearcă să treacă la o stare de echilibru, adică este complementar cu primul postulat.
Apare o întrebare rezonabilă: dacă în orice moment natura implementează numai stări și procese optime, de ce există atâtea absurdități și erori în lume care sunt departe de conceptul de optimitate? Sau poate persoana care a condus planeta la un dezastru ecologic este o excepție pentru care legea optimității nu este scrisă? Dar nu numai oamenii comit acte absurde. Există vreo optimitate în comportamentul unei molii care zboară spre un incendiu, sau un roi de lăcuste care distrug toată vegetația din zonă și apoi moare de foame, sau o muscă care bate de sticlă? Se pare că există. De exemplu, o muscă care lovește paharul folosește unul dintre cei mai eficienți algoritmi pentru găsirea soluției optime: metoda de căutare aleatorie. O muscă nu are aparatul analitic pe care îl are o persoană. Înțelegem că trebuie să vă abateți ușor în lateral și să zburați pe fereastra deschisă. Musca nu știe dacă există o cale de ieșire din situația în care se află. Dar o căutare aleatorie garantează că o soluție va fi găsită mai devreme sau mai târziu, dacă este posibil în principiu. Mai mult decât atât, o căutare aleatorie ne permite uneori să găsim o cale de ieșire chiar și din situații aparent fără fund (așa își poate găsi propria soluție la o problemă, și nu cea pe care am pregătit-o pentru aceasta, de exemplu, poate găsim şi zburam într-un gol de care nici nu ştiam).nu bănuiam). În plus, dacă funcția (situația) în sine este aleatorie în natură (sau natura ei este necunoscută), atunci această metodă oferă cel mai scurt timp mediu pentru căutarea extremului său.
Natura folosește foarte des astfel de algoritmi de optimizare. Deci, de exemplu, tacticile de căutare a locurilor de mită (nectar și polen) efectuate de o colonie de albine sunt foarte indicative în acest sens. Dacă una dintre albine găsește o poiană bogată de flori, atunci la întoarcerea în stup, ea își execută faimosul „dans pe fagure”, care „spune” celorlalte albine unde să zboare, câtă energie va necesita acest lucru, ce fel de flori. cresc în poiană etc. P. După aceasta, multe albine zboară la destinație. În același timp, ei demonstrează o bună înțelegere a informațiilor care le sunt transmise. Dar din anumite motive, nu toate albinele care au urmărit dansul sunt suficient de punctuali.
Unii dintre ei se rătăcesc sau chiar zboară inițial în direcția greșită, uneori în direcția complet opusă. Acest lucru reduce cantitatea de mită adusă în stup. Dar se dovedește că astfel de erori sunt programate inițial și sunt de mare beneficiu. În principiu, natura ar putea recompensa albinele cu o infailibilitate absolută asemănătoare unui robot în a se înțelege reciproc. Dar ea le-a dat albinelor „dreptul de a greși”. Chiar și procentul de albine care s-au rătăcit este determinat destul de strict (aproximativ 5%). Zborurile „eronate” aduc de obicei în stup informații despre alte surse bogate de mită, de care aceste albine se poticnesc uneori accidental.
Fără o anumită cantitate de eroare, absurditate și șansă, natura nu și-ar putea dezvolta și complica formele. De aceea, probabil, A.S. Pușkin a numit cazul „zeu-inventator”. Aici se realizează însăși libertatea de alegere, fără de care armonia în sistem este de neconceput. Sistemele a căror structură este lipsită de incertitudine, aleatoriu și eroare sunt neviabile, deoarece sunt departe de armonie - starea optimă cea mai obiectivă. Ele nu se pot dezvolta și, prin urmare, sunt inutile pentru Univers. Prin urmare, ele sunt distruse destul de repede (eroare acumulată). Mecanismele pentru aceasta sunt variate. Relația armonioasă dintre predeterminarea strictă și libertatea de alegere în structura sistemului este determinată de „proporția de aur”.
- 3.Izolarea biologică și rolul acesteia în transformarea populațiilor.
Izolarea (din franceză izolare - separare, separare) (biologică), restricție sau încălcare a încrucișării libere a indivizilor și amestecarea diferitelor forme de organisme; unul dintre factorii elementari ai evoluţiei. Charles Darwin, folosind exemplul faunei și florei insulare, a arătat rolul alimentelor în apariția, expansiunea și adâncirea diferențelor dintre forme similare de organisme vii. Dacă vreuna, adesea periferică, o parte a populației inițiale este izolată de unele bariere geografice, atunci în timp această parte a populației se poate transforma într-o specie independentă. Această metodă geografică de speciație, potrivit multor biologi, este singura sau, în orice caz, principala modalitate de speciație.
În termeni macroevoluționari, I. este determinat de neîncrucișarea diferitelor specii, adică are predominant caracterul de I reproductor. În termeni microevoluționari, adică la nivel intraspecific, se disting două grupe principale de I.: teritorial-mecanici. , care include toate cazurile de apariție a barierelor între diferite părți ale populației sau diferite populații (de exemplu, bariere de apă pentru pământ și pământ pentru organismele acvatice, munți pentru văi și văi pentru speciile montane etc.), și biologice, care este împărțit în 3 subgrupe:
a) ecologic I. - indivizii a două sau mai multe biotipuri se găsesc rar sau deloc în timpul perioadei de reproducere:
b) morfo-fiziologic I. - copularea este dificilă sau imposibilă din motive morfologice sau etologice (comportamentale);
c) I. genetic propriu-zis, cauzat de inferioritatea (viabilitate redusă, fertilitate sau sterilitate completă) a hibrizilor obținuți ca urmare a încrucișărilor adecvate.
Toate tipurile de I. pot exercita presiuni diferite asupra populațiilor, deoarece orice formă de I. poate fi exprimată cantitativ în grade diferite. I. teritorial-mecanic (în arii extinse – geografice) duce la morfogeneză alopatrică și, cu acțiune suficient de lungă, determină de obicei apariția unei forme de I biologic. Cazurile de apariție primară a I. biologice pot duce la morfogeneză simpatrică.
Izolarea biologică este asigurată de două grupe de mecanisme: cele de eliminare a încrucișării (pre-copulatorii) și izolarea în timpul încrucișării (post-copulatorii).
Împerecherea formelor strâns înrudite este împiedicată de diferențele din timpul activității sexuale și de maturarea produselor reproductive. În natură, izolarea biotopică este comună, în care potențialii parteneri de împerechere nu se întâlnesc, deoarece trăiesc adesea în locuri diferite. Astfel, unele cinteze (Fringillacoelebs) cuibăresc în regiunea Moscovei în păduri de tip taiga, iar altele în arborete joase și rare, cu un număr mare de poieni. Potențialul de împerechere încrucișată a indivizilor acestor grupuri este oarecum limitat. Un exemplu interesant de izolare biotopică îl reprezintă formele intraspecifice simpatrice la cucul comun (Cuculuscanorus). Europa găzduiește mai multe „rase biologice” de cuci, care diferă prin culoarea determinată genetic a ouălor lor. În Europa de Est, unii depun ouă albastre în cuiburile pufului roșu comun și ale stonechat, alții depun ouă pete deschise în cuiburile micilor păsări paseriforme care au ouă de o culoare similară. Izolarea între aceste forme de cuci este menținută prin distrugerea ouălor insuficient camuflate de către speciile gazdă. La multe specii, preferința biotopului este un mecanism eficient de izolare.
De o mare importanță în apariția și menținerea izolării biologice în forme strâns înrudite este izolarea etologică - complicații ale împerecherii cauzate de caracteristicile comportamentale. La prima vedere, diferențele nesemnificative în ritualul de curte și schimbul de stimuli vizuali, sonori și chimici vor împiedica continuarea curtației.
Un mecanism de izolare important care face dificilă încrucișarea speciilor strâns înrudite este apariția diferențelor morfofiziologice în organele reproducătoare (izolare morfofiziologică).
Al doilea grup mare de mecanisme de izolare din natură este asociat cu apariția izolării după fertilizare (izolare genetică intrinsecă), inclusiv moartea zigoților după fertilizare, dezvoltarea hibrizilor complet sau parțial sterili, precum și viabilitatea redusă a hibrizilor.
- 4.Imaginația geno- și fenotip.
Genotip, totalitatea tuturor genelor localizate în cromozomii unui organism dat. Într-un sens mai larg, genetica este totalitatea tuturor factorilor ereditari ai unui organism - atât nucleari (genomul), cât și nenuclearii, extracromozomiali (adică factorii ereditari citoplasmatici și plastici). Termenul a fost propus de biologul danez V. Johansen (1909). G. este un purtător de informații ereditare transmise din generație în generație. Este un sistem care controlează dezvoltarea, structura și activitatea vitală a unui organism, adică totalitatea tuturor caracteristicilor unui organism - fenotipul acestuia. G. este un sistem unificat de gene care interacționează, astfel încât manifestarea fiecărei gene depinde de mediul genotipic în care se află. De exemplu, culoarea roșie a florilor la unele soiuri de mazăre dulce apare numai cu prezența simultană a alelelor dominante a două gene diferite în G., în timp ce separat fiecare dintre aceste alele provoacă culoarea albă a florilor. Interacţiunea lui G. cu un complex de factori din mediul intern şi extern al organismului determină manifestarea fenotipică a simptomelor. Un exemplu de influență a mediului asupra manifestării fenotipice a lui G. este culoarea blănii iepurilor, așa-numita. Linia himalayană: la aceeași temperatură, acești iepuri, când sunt crescuți la frig, au blana neagră, la temperaturi moderate - colorare „himalayană” (alb cu bot, urechi, labe și coadă negru), la temperaturi ridicate - blană albă. Descendenții acestor trei grupuri de animale nu moștenesc o culoare neschimbată a blanii, ci capacitatea de a produce o anumită culoare, diferită în diferite condiții de mediu. Prin urmare, în termeni generali, este mai corect să spunem că genetica determină moștenirea nu a unor trăsături specifice, ci a normei de reacție a organismului și a tuturor condițiilor posibile de mediu. În diferite stadii de dezvoltare a unui individ, uneori una sau alte gene sunt în stare activă; prin urmare, în ontogeneză G. funcţionează ca un sistem mobil variabil.
Termenul „G”. folosit uneori într-un sens mai restrâns pentru a se referi doar la un grup de gene sau chiar la gene individuale, a căror moștenire este subiect de observație.
De exemplu, în descendenții de scindare dintr-o încrucișare monohibridă AA (aa), se obișnuiește să se vorbească despre genotipurile AA, Aa și aa, făcând abstracție de la posibilele diferențe între indivizii (sau grupurile de indivizi) corespunzători din alte gene.
Fenotipul, caracteristicile structurale și funcțiile vitale ale unui organism, determinate de interacțiunea genotipului acestuia cu condițiile de mediu. În sens larg, termenul „F.”, propus de D. biologul V. Johansen în 1909, denotă întregul set de manifestări ale genotipului (aspectul general al organismului), iar în sens restrâns - caracteristici individuale (fene) controlate de anumite gene. Conceptul de fiziologie se extinde la orice caracteristică a unui organism, pornind de la produsele primare ale acțiunii genelor - molecule de ARN și polipeptide și terminând cu caracteristici ale structurii externe, proceselor fiziologice, comportamentului etc.
La nivelul produselor primare ale acțiunii genelor, legătura dintre genotipul unui organism și fizica acestuia este destul de clară: fiecare secvență de nucleotide dintr-o moleculă de acid dezoxiribonucleic (ADN) corespunde unei secvențe bine definite de nucleotide dintr-un ribonucleic. moleculă de acid (ARN) și, în consecință, o secvență specifică de aminoacizi dintr-un lanț polipeptidic (protein). Totuși, chiar și la acest nivel, secvența nucleotidelor din ADN, adică structura sa primară, determină fără ambiguitate doar structura ARN-ului și proteinelor sintetizate pe baza acestuia, dar nu și momentul sintezei lor sau cantitatea acestor produse, la fel ca o matrice tipografică determină conținutul textului, dar nu și momentul tipăririi sau difuzării acestuia. Timpul de activare al genelor individuale și intensitatea „lecturii” acestora depind atât de munca anterioară a altor gene, cât și de un complex de factori intracelulari și de mediu.
La niveluri superioare de organizare biologică, adică la nivelul celulelor, țesuturilor, organelor, sistemelor de organe și organismului în ansamblu, relația dintre genotip și F. este și mai complexă. În aceste cazuri, fiecare trăsătură este rezultatul interacțiunii produselor multor gene, care, la rândul lor, depinde de condițiile specifice de mediu. Acest lucru se vede mai ales în mod clar în exemplul caracteristicilor cantitative. Studiind greutatea boabelor în mai multe linii de plante autopolenizate, adică omogene genetic, de plante de fasole, Johansen a descoperit că la plantele de același genotip, boabele variază în greutate, adică au F diferite. Mai mult decât atât, limitele variabilității în greutatea boabelor de linii diferite se suprapun adesea (unul și același F. poate corespunde unor genotipuri diferite). În același timp, între diferite linii crescute în aceleași condiții, există diferențe ereditare stabile în greutatea medie a boabelor. Cu toate acestea, greutatea medie a boabelor din fiecare linie poate varia în funcție de condițiile de mediu, de exemplu, nutriția plantelor. Astfel, rolul relativ al eredității și al mediului în dezvoltarea trăsăturilor poate fi foarte diferit. Studiul trăsăturilor cantitative care sunt de mare importanță practică în agricultură și medicină se realizează folosind metode biometrice speciale pentru analizarea eredității trăsăturilor.
- 5. Poluarea solului.
Poluarea solului este cauzată de o varietate de substanțe - microelemente de metale, microdoze de poluanți organici, produse de eliminare și dezinfectare a apelor uzate, produse de protecție a plantelor, hidrocarburi și substanțe radioactive. Poluarea solului are un efect dăunător asupra plantelor, ducând la acumularea de elemente toxice în ele; această bioacumulare este periculoasă și pentru oameni. În plus, substanțele chimice care intră în pământ pot provoca coroziunea utilităților subterane. Cele mai contaminate soluri sunt în zonele miniere, în locurile de agricultură intensivă, precum și solurile din pajişti și terenuri arabile adiacente drumurilor, terenuri în care se deversează apele uzate de la întreprinderile agroindustriale sau nămolurile formate în urma epurării apelor uzate urbane. Majoritatea terenurilor contaminate sunt situate în mari regiuni industriale. Poluarea tehnogenă a solului a necesitat dezvoltarea unor metode speciale de regenerare și protecție a acestuia.
Poluarea chimică a solului este o modificare a compoziției chimice a solului care a apărut sub influența directă sau indirectă a factorilor de utilizare a terenurilor (industriali, agricoli, municipali), determinând o scădere a calității acestuia și un posibil pericol pentru sănătatea publică.
Poluarea biologică a solului este parte integrantă a poluării organice cauzată de diseminarea (răspândirea) agenților patogeni ai bolilor infecțioase și invazive, precum și a insectelor și acarienilor dăunători, purtători de agenți patogeni ai bolilor umane, animale și plante.
- 6. Ploaia acidă, originea ei, consecințele impactului asupra obiectelor naturale.
Termenul „ploaie acidă” a fost inventat pentru prima dată în acest an de către cercetătorul englez Angus Smith. Smogul victorian din Manchester i-a atras atenția. Și deși oamenii de știință din acea vreme au respins teoria existenței ploii acide, astăzi nimeni nu se îndoiește că ploaia acide este una dintre cauzele morții vieții în corpurile de apă, păduri, culturi și vegetație. În plus, ploaia acide distruge clădirile și monumentele culturale, conductele, face mașinile inutilizabile, reduce fertilitatea solului și poate duce la scurgerea de metale toxice în acvifere.
Apa ploii obișnuite este, de asemenea, o soluție ușor acidă. Acest lucru se întâmplă deoarece substanțele atmosferice naturale, cum ar fi dioxidul de carbon (CO2), reacționează cu apa de ploaie. În acest caz, se formează acid carbonic slab (CO2 + H2O -> H2CO3). . În timp ce pH-ul ideal al apei de ploaie este de 5,6-5,7, în viața reală aciditatea (pH) a apei de ploaie dintr-o zonă poate diferi de aciditatea apei de ploaie dintr-o altă zonă. Acest lucru, în primul rând, depinde de compoziția gazelor conținute în atmosfera unei anumite zone, cum ar fi oxidul de sulf și oxizii de azot.
În 2009, omul de știință suedez Svante Arrhenius a inventat doi termeni - acid și bază. El a numit acizi substanțe care, atunci când sunt dizolvate în apă, formează ioni de hidrogen liber încărcați pozitiv (H+). El a numit baze substanțe care, atunci când sunt dizolvate în apă, formează ioni de hidroxid liber încărcați negativ (OH-). Termenul pH este folosit ca indicator al acidității apei. Termenul pH înseamnă, tradus din engleză, un indicator al gradului de concentrație a ionilor de hidrogen.
Valoarea pH-ului este măsurată pe o scară de la 0 la 14. Apa și soluțiile apoase conțin atât ioni de hidrogen (H+) cât și ioni de hidroxid (OH). Când concentrația ionilor de hidrogen (H+) în apă sau soluție este egală cu concentrația ionilor de hidroxid (OH-) din aceeași soluție, atunci o astfel de soluție este neutră. Valoarea pH-ului unei soluții neutre este 7 (pe o scară de la 0 la 14). Când acizii sunt dizolvați în apă, crește concentrația ionilor liberi de hidrogen (H+). De asemenea, cresc aciditatea apei. În același timp, odată cu creșterea concentrației ionilor de hidrogen (H+), concentrația ionilor de hidroxid (OH-) scade. Acele soluții a căror valoare pH pe scara dată variază de la 0 la<7, называются кислыми. Когда в воду попадают щелочи, то в воде повышается концентрация гидроксид-ионов (ОН-). При этом в растворе понижается концентрация ионов водорода (Н+). Растворы, значение рН которых находится в пределах от >7 până la 14 sunt numite alcaline.
Trebuie remarcată încă o caracteristică a scalei pH. Fiecare etapă ulterioară pe scara pH indică o scădere de zece ori a concentrației ionilor de hidrogen (H+) (și, în consecință, aciditatea) în soluție și o creștere a concentrației ionilor de hidroxid (OH-). De exemplu, aciditatea unei substanțe cu o valoare a pH-ului de zece ori mai mare decât aciditatea unei substanțe cu o valoare a pH-ului de 5, de o sută de ori mai mare decât aciditatea unei substanțe cu o valoare a pH-ului de 6 și o sută de mii. de ori mai mare decât aciditatea unei substanțe cu valoarea pH-ului 9.
Ploaia acidă se formează printr-o reacție între apă și poluanți precum oxidul de sulf (SO2) și diverși oxizi de azot (NOx). Aceste substanțe sunt emise în atmosferă prin transportul rutier, ca urmare a activităților întreprinderilor metalurgice și centralelor electrice.
Compuși ai sulfului (- H2S (în cantități mici, cu ardere insuficientă sau ardere incompletă, la temperaturi scăzute). Diferiți compuși ai azotului sunt conținuți în cărbuni, și mai ales în turbă (deoarece azotul, ca și sulful, face parte din structurile biologice, din care aceștia s-au format minerale).Când astfel de minerale sunt arse, se formează oxizi de azot (oxizi acizi, anhidride) - de exemplu, oxid de azot (IV) NO2. Reacționând cu apa atmosferică, se transformă în soluții de acizi - sulfuric, sulfuros, azotos și nitric.Apoi, odata cu zapada sau ploaia, cad la pamant.
Consecințele ploii acide sunt observate în SUA, Germania, Cehia, Slovacia, Țările de Jos, Elveția, Australia, republicile fostei Iugoslavii și multe alte țări de pe glob.
Ploaia acidă are un impact negativ asupra corpurilor de apă - lacuri, râuri, golfuri, iazuri - crescându-le aciditatea la un asemenea nivel încât flora și fauna mor în ele. Plantele acvatice cresc cel mai bine în apă cu valori ale pH-ului între 7 și 9,2. Odată cu creșterea acidității (valorile pH-ului se deplasează la stânga punctului de referință 7), plantele acvatice încep să moară, privând alte animale de rezervor de hrană. La aciditate pH 6, creveții de apă dulce mor. Când aciditatea crește la pH 5,5, bacteriile de la fund care descompun materia organică și frunze mor, iar resturile organice încep să se acumuleze în partea de jos. Apoi moare planctonul - un animal minuscul care formează baza lanțului trofic al rezervorului și se hrănește cu substanțele formate atunci când bacteriile descompun substanțele organice. Când aciditatea atinge pH-ul 4,5, toți peștii, majoritatea broaștelor și insectelor mor.
Pe măsură ce materia organică se acumulează pe fundul corpurilor de apă, metalele toxice încep să se scurgă. Aciditatea crescută a apei promovează o solubilitate mai mare a metalelor periculoase, cum ar fi aluminiul, cadmiul și plumbul din sedimente și sol.
Aceste metale toxice prezintă un risc pentru sănătatea umană. Persoanele care beau apă cu un nivel ridicat de plumb sau mănâncă pește cu un nivel ridicat de mercur se pot îmbolnăvi grav.
Ploaia acidă dăunează mai mult decât doar vieții acvatice. De asemenea, distruge vegetația de pe uscat. Oamenii de știință cred că, deși mecanismul nu este încă pe deplin înțeles, „un amestec complex de poluanți, inclusiv precipitații acide, ozon și metale grele, duc în mod colectiv la degradarea pădurilor.
Pierderile economice din cauza ploilor acide în SUA sunt estimate de un studiu la 13 milioane de dolari anual pe Coasta de Est, iar până la sfârșitul secolului pierderile vor ajunge la 1,750 de miliarde de dolari din pierderea pădurilor; 8,300 miliarde dolari în pierderi de recoltă (numai în bazinul râului Ohio) și 40 milioane dolari în cheltuieli medicale numai în Minnesota. Singura modalitate de a schimba situația în bine, potrivit multor experți, este reducerea cantității de emisii nocive în atmosferă.
- 7.Tehnologii pentru reciclarea deșeurilor industriale.
Necesitatea asigurării siguranței mediului și a creșterii eficienței economice a eliminării deșeurilor a forțat oamenii de știință și specialiștii să dezvolte tehnologia casnică pentru reciclarea la temperatură înaltă (1500-1600°C) a deșeurilor solide municipale cu producerea de gaz de piroliză și generarea de energie electrică. Această tehnologie face posibilă procesarea deșeurilor cu orice putere calorică și umiditate; concomitent cu deșeurile solide, este posibilă procesarea unor tipuri de deșeuri industriale. Piroliza la temperatură înaltă a gazelor cu descompunerea completă a componentelor organice și purificarea lor permite arderea lor ulterioară în cazane industriale. Energia electrică rezultată pentru nevoile proprii și vânzarea către consumatori este de 2 ori mai ieftină decât cea obținută prin metode tradiționale.
Tehnologie ecologică pentru reciclarea deșeurilor lichide de la întreprinderile miniere. Tehnologia de tratare a apelor uzate se bazează pe extracția prin chimisorbție a ionilor metalici grei și rari folosind materiale filtrante polimerice VION și include următoarele etape:
1) filtrare și sedimentare preliminară pentru a îndepărta impuritățile mecanice din soluțiile apoase;
2) purificarea prin sorbție din ioni Te (II), Te (III), Co (II) și Cu (II) cu schimbător de cationi KN-1 și din ionii W (VI) și Mo (VI) cu anion AS-1 în dinamică mod la pH Miercuri 6-8;
3) regenerarea separată acido-bazică a filtrelor schimbătoare de cationi și anioni cu concentrația ionilor desorbiți în eluați;
4) utilizarea reactivilor de sodă ca produse industriale în schema principală a procesului tehnologic;
5) utilizarea apei purificate într-un sistem închis de circulație a apei.
Instalatii mobile pentru prelucrarea deseurilor radioactive lichide. Scopul unităților mobile: prelucrarea deșeurilor radioactive lichide de nivel scăzut în scopul epurării apei la standardele de siguranță împotriva radiațiilor, concentrarea impurităților toxice într-un volum mic pentru a reduce cantitatea totală de deșeuri radioactive.
Instalațiile Întreprinderii Unitare de Stat MosNPO „Radon” utilizează procesele de filtrare, sorbție, ultrafiltrare, osmoză inversă, electrodializă, coagulare și electroosmoză.