Репарация как механизм поддержаня генетического гомеостаза.виды и механизмы репарации. Системы репарации ДНК: общие сведения Варианты репарации днк
Принципы репарации ДНК у различных организмов сходны. Ряд повреждений клетка удаляет из ДНК путем прямой реактивации. Таким образом исправляются алкилированные азотистые основания. К этому же типу репарации относится и удаление тиминовых димеров на свету. Другие виды репарации ультрафиолетовых повреждений ДНК называют темновой репарацией, чтобы отличить от прямой фотореактивации.
Если невозможна прямая реактивация, работают механизмы эксцизионной репарации , удаляющие из ДНК нарушенные участки. При этом типе репарации специальные эндонуклеазы производят разрез одной цепи ДНК вблизи места повреждения. Далее экзонуклеазы удаляют поврежденный участок. Образовавшуюся брешь заполняет ДНК-полимераза, а остающийся разрыв сшивает ДНК-лигаза. Видно, что эксцизионная репарация всегда использует один и тот же принцип: нарушенный участок ДНК удаляется, а затем восстанавливается на матрице ненарушенной комплементарной цепи ДНК.
Индуцируемая репарация. В условиях, увеличивающих количество повреждений ДНК, происходит индукция дополнительных репаративных ресурсов клетки. У бактерий индуцируемая репарация используется лишь в тех случаях, когда повреждений в ДНК становится настолько много, что это начинает угрожать клетке гибелью. Поэтому индуцируемая система репарации называется SOS-репарацией . Степень индукции SOS-системы определяется количеством повреждений. Степень индукции SOS-системы в определенном смысле отражает «благополучие» клетки и ее шансы на выживание. Поэтому некоторые умеренные бактериофаги используют индукцию SOS-системы в качестве сигнала для размножения и уничтожения клетки-хозяина.
Дублирование информации в двух комплементарных цепях ДНК не позволяет безошибочно исправлять все типы повреждений. Описанные механизмы репарации не могут справиться с такими нарушениями структуры ДНК, как ковалентные межнитевые сшивки, которые могут возникать под действием ряда мутагенов, или двухцепочечные разрывы ДНК. Такие повреждения могут репарироваться только при наличии гомологичной неповрежденной молекулы ДНК, т.е. рекомбинационным путем.
7608 0
Под регенерацией подразумевают восстановление тканью, органом утраченной или поврежденной специализированной структуры.
Физиологическая регенерация заключается в обновлении морфофункциональных свойств ткани или органа с помощью естественных механизмов, например, образовании новых и резорбции старых, изношенных остеонов в кости.
При репаративной регенерации происходит процесс формирования новых структур на месте повреждения или травмы. В качестве иллюстрации можно привести процесс перелома длинных трубчатых костей. Процессы репаративной регенерации клеток, заключающиеся в образовании тканей на месте гибели поврежденных элементов во многом регулируются механическими условиями. В частности, деформация регенерата, например растяжение, из-за нестабильности может стимулировать как образование костной мозоли, так и рассасывание кости в зоне контактирующих поверхностей. Если происходит рассасывание, то возрастает нестабильность в зоне перелома. Увеличивающаяся деформация регенерата, например с использованием компрессионно-дистракционных аппаратов для остеосинтеза, может привести к постепенной дифференцировке клеток стромы в сторону повышения их прочности и жесткости. Так, мягкая грануляционная ткань, способная выдержать существенную деформацию, замещается соединительной тканью, обладающей большей жесткостью, но меньшей прочностью к деформации. Этот процесс часто называют «непрямым» заживлением. Если щель перелома небольшая и костные отломки хорошо стабилизированы межфрагментарной компрессией, то деформация проявляется минимально. При этом часто возникает прямое образование костной ткани, а рассасывание кости и образование периостальной мозоли не всегда обязательно. Такой тип заживления переломов называют «прямым» (контактным).
После санации очага воспаления от микробных и чужеродных тел включаются механизмы, протекающие с участием лимфоцитов и макрофагов. Лимфоциты секретируют ИЛ-2 и ФНО, которые активируют моноциты крови, которые в тканях проходят через стадию примирования и трансформируются в активированные макрофаги. Эти клетки, в свою очередь, секретируют в окружающую ткань ростовые факторы типа ФРФ, тромбоцитарный фактор роста, ИЛ-6, которые оказывают влияние на остеобласты, фибробласты и эндотелиальные клетки. Фибробласты делятся и по мере созревания начинают секретировать компоненты экстрацеллюлярного матрикса (протеогликаны, гликозаминогликаны, фибронектин, адгезины и т.п.), включая коллаген. Макрофаги контролируют фибриллогенез путем продукции при необходимости ферментов - коллагеназы и эластазы. Следует отметить, что оптимум работы большинства изо-форм этих ферментов лежит в нейтральной среде, т.е. тогда, когда все кислые продукты в очаге воспаления уже удалены или нейтрализованы. Кроме того, макрофаги через секрецию простагландинов и ФРФ, ФРТ и других факторов могут стимулировать или супрессировать функцию фибробластов, оказывая тем самым влияние на объем новой ткани (Кетлицкий, 1995; Серов и др., 1995).
Параллельно активируются процессы ангиогенеза. При этом макрофаги как бы пробивают туннели в экстрацеллюлярном матриксе, в которые мигрируют клетки эндотелия. При этом возникают новые капилляры, которые растут, превращаются в более крупные сосуды, ветвятся и пронизывают новую ткань (Маянский, Урсов, 1997). Этот процесс в какой-то мере напоминает механизм аппозиционного роста костной ткани или образования костной мозоли при переломах, в котором прослеживается та же, по-видимому, общебиологическая последовательность событий.
В результате заживления раны образуется новая ткань, которая в той или иной мере замещает функцию поврежденных структур. К сожалению, не всякое воспаление заканчивается таким исходом. В ряде случаев оно происходит с образованием разнообразных дефектов, грубой рубцовой тканью, переходит в хроническую стадию, включает аутоиммунные механизмы и склерозирование (кальцификации) тканей.
А.В. Карпов, В.П. Шахов
Системы внешней фиксации и регуляторные механизмы оптимальной биомеханики
История открытия
Однонитевое и двунитевое повреждения ДНК
Источники повреждения ДНК
- УФ излучение
- Химические вещества
- Ошибки репликации ДНК
- Апуринизация - отщепление азотистых оснований от сахарофосфатного остова
- Дезаминирование - отщепление аминогруппы от азотистого основания
Основные типы повреждения ДНК
- Повреждение одиночных нуклеотидов
- Повреждение пары нуклеотидов
- Разрыв цепи ДНК
- Образование поперечных сшивок между основаниями одной цепи или разных цепей ДНК
Устройство системы репарации
Каждая из систем репарации включает следующие компоненты:
- фермент , "узнающий" химически изменённые участки в цепи ДНК и осуществляющий разрыв цепи вблизи от повреждения
- фермент , удаляющий повреждённый участок
- фермент (ДНК-полимераза), синтезирующий соответствующий участок цепи ДНК взамен удалённого
- фермент (ДНК-лигаза), замыкающий последнюю связь в полимерной цепи и тем самым восстанавливающий её непрерывность
Типы репарации
Эксцизионная репарация
Эксцизионная репарация (англ. excision - вырезание) включает удаление повреждённых азотистых оснований из ДНК и последующее восстановление нормальной структуры молекулы.
Примечания
Wikimedia Foundation . 2010 .
Смотреть что такое "Репарация (биология)" в других словарях:
Система обнаружения и репарации вставок, пропусков и ошибочных спариваний нуклеотидов, возникающих в процессе репликации и рекомбинации ДНК, а также в результате некоторых типов повреждений ДНК Сам факт ошибочного спаривания не позволяет… … Википедия
У этого термина существуют и другие значения, см. Репарация. Повреждённые хромосомы Репарация особая функция клеток, заключающаяся в способности исправлять химические повреждения и разрывы в молекулах ДНК, повреждённой при нормальном… … Википедия
Радиационная биология или радиобиология наука, изучающая действие ионизирующих и неионизирующих излучений на биологические объекты. Код науки по 4 х значной классификации ЮНЕСКО (англ.) 2418 (раздел биология). Радиобиология … Википедия
I Генетика (от греч. génesis происхождение) наука о законах наследственности и изменчивости организмов. Важнейшая задача Г. разработка методов управления Наследственностью и наследственной Изменчивостью для получения нужных человеку форм… … Большая советская энциклопедия
Специализация: Клеточная биология Периодичность: ежемесячно Сокращённое название: Nat. Cell. Biol. Язык: Английский Главный редактор: Саумья Суомината … Википедия
Мутагенез это внесение изменений в нуклеотидную последовательность ДНК (мутаций). Различают естественный (спонтанный) и искусственный (индуцированный) мутагенез. Содержание 1 Естественный мутагенез … Википедия
Радиационная биология или радиобиология это самостоятельная комплексная, фундаментальная наука, состоящая из многих научных направлений, изучающая действие ионизирующих и неионизирующих излучений на биологические объекты. Код науки по 4 х значной … Википедия
Halobacteria, штамм NRC 1, каждая клетка около 5 мкм длиной … Википедия
Репарация ДНК
Общие сведения
Агенты повреждающие ДНК
Излучение
1. ионизирующая радиация (гамма лучи, рентгеновские лучи)
2. Ультрафиолетовое излучение (особенно ~260 нм, именно в этой области происходит максимальное поглощение ДНК)
Активные радикалы кислорода образуемые во время нормального клеточного дыхания в различных биохимических путях.
Химические вещества окружающей среды.
Многие углеводороды.
Химикаты используемые в противоопухолевой химотерапии.
Типы повреждений ДНК
1. Все четыре основания в ДНК (A, T, C, G) могут быть ковалентно модифицированы в различных положениях.
Наиболее частое - это потеря аминогруппы (дезаминирование) - в таком случае C превращается в U.
Неправильное включение оснований происходящее из-за ошибок в работе ДНК полимераз при репликации.
Наиболее часто происходит включение урацила вместо тимина.
Нарушения структуры.
В ДНК могут происходить разрывы. Разрывы могут быть одноцепочечные, или могут разорваться обе цепи ДНК.
Ионизирующая радиация может быть частой причиной таких разрывов.
Между соседними основаниями может образоваться ковалентная связь, причем связь может образоваться между соседними основаниями в одной цепи или между двумя цепями ДНК.
типы повреждения ДНК
изменение одного основания
апуринизация
замена С на У
замена А на гипоксантин
алкилирование основания
вставка или делеция нуклеотида
встраивание аналогичного основания
изменение двух оснований
образование тиминовых димеров
поперечная связь с бифункиональным алкилирующим агентом
разрушение цепи
ионизирующее излучение
радиоактивное разрушение элементов остова
поперечные связи
между основаниями одной нити или двух параллельных нитей
между ДНК и белковыми молекулами, например гистонами
Репарация поврежденных оснований
Поврежденные основания могут быть исправлены различными путями:
Прямое химическое исправление повреждений.
Эксцизионная репарация (ER), при которой поврежденное основание удаляется и заменяется новым. Имеется три модели эксцизионной репарации, в каждой из которых используется свой собственный набор ферментов.
Эксцизионная репарация оснований (BER).
Эксцизионная репарация нуклеотидов (NER).
Мисмэтч репарация(MMR).
Прямое исправление повреждений.
Наиболее частая причина точечных мутаций у человека - это спонтанное добавление метильной группы - один из типов алкилирования. Такие модификации исправляются ферментами называемыми гликозилазами, исправляющими ошибку без разрушения цепи ДНК.
Некоторые препараты используемые в химотерапии также повреждают ДНК путем алкилирования.
Проблема репарации состоит в том, что ограниченным набором ферментов и механизмов клетка должна справиться со многими повреждениями вызванными самыми различными химическими и физическими агентами.
Эксцизионная репарация оснований (BER)
Основные ключевые события:
1. Удаления поврежденного основания (происходит ~ 20,000 раз в день в каждой клетке человеческого тела) ДНК гликозилазими. У человека имеется по крайней мере 8 генов кодирующих различные ДНК гликозилазы, кадждые из которых распознают свой набор повреждений оснований.
2. Удаление дезоксирибофосфата приводит к образованию пустоты в ДНК.
3. Замена правильным нуклеотидом. Это функция у человека выполняется ДНК полимеразой бетта.
4. Лигирование разрыва цепи. Имеется два фермента, оба нуждаются в АТФ.
Эксцизионная репарация нуклеотидов (NER)
NER отличается от BER несколькими путями.
Использованием различных ферментных систем.
Даже если ошибка в одном нуклеотиде, удаляется сразу множество нуклеотидов в районе повреждения.
Основные ключевые события NER:
1.Повреждение распознается одним или несколькими факторами связывающимися с местом повреждения.
2. ДНК раскручивается в месте повреждения. В этом процессе участвуют
различные транскрипционные факторы IIH, TFIIH, (которые так же работают при нормальной транскрипции).
3. Разрез ДНК происходит с 3" и 5"-конца от повреждения, в результате чего удаляется фрагмент ДНК содержащий поврежденный нуклеотид.
4. Новая цепь ДНК достраивается по матрице неповрежденной цепи ДНК полимеразами дельта или эпсилон.
5. Лигазы сшивают вновь синтезированный конец цепи.
Пигментная ксеродерма (XP)
XP редкое наследственное заболевание человека проявляющееся в поражении кожи при облучении светом, что в конечном итоге приводит к развитию рака кожи и гибели больного.
Болезнь возникает из-за мутаций в генах участвующих в NER репарации. Например:
XPA кодирует белок связывающийся с местом повреждения и помогающий сборке репарирующего комплекса.
XPB и XPD, которыя являются частями транскрипционного фактора TFIIH. Некоторые мутации в XPB и XPD также могут являться причиной преждевременного старения.
XPF разрезает цепь ДНК с 5"-конца от повреждения.
XPG разрезает цепь с 3"-конца.
Мисмэтч репарация(MMR)
Мисмэтч репарация исправляет ошибочно встроенные неповрежденные основания которые не образуют нормальное Уотсон-Криковское спаривание (A T, C G). Такие ошибки происходят во время работы ДНК полимеразы при репликации.
В мисмэтч репарации участвуют ферменты вовлеченные как в BER, так и в NER репарацию, так и специализированные ферменты.
Синтез ДНК при мисмэтч репарации осуществляется ДНК полимеразами дельта или эпсилон.
Система мисмэтч репарации участвует в увеличении точности рекомбинации при мейозе.
Репарация разрывов ДНК
Ионизирующая радиация и некоторые химические вещества способны разорвать одну или две цепи ДНК.
Одноцепочечные разрывы (SSB)
Разрывы одной из цепей ДНК часто исправляются ферментами участвующими в BER репарации.
Двуцепочечные разрывы (DSB)
Имеется два механизма которые способны устранить двуцепочечных разрывов ДНК:
Прямое соединение сломаных концов. Этот процесс требует специальных
ферментов, которые узнают и связывают разорванные концы с последующим их сшиванием. Если разорванная ДНК имеет тупые концы и соединение двух фрагментов ДНК происходит случайно, то такая репарация называется NHEJ. Белок Ku необходимый для NHEJ. Ku - гетеродимерная субъединица, состоящая из двух белков Ku70 и Ku80.
Ошибки возникающие при прямом присоединении могут являться причиной транслокаций.
Полинуклеотидлигаза
– восстановл. одноцеп. разрывы ДНК
Гомологичная рекомбинация
Гомологичная рекомбинация способна восстанавливать поломанные концы хромосом используя ДНК не поврежденной сестринской хроматиды доступной после удвоения хромосом.
Гены необходимые для гомологичной рекомбинации - BRCA-1 and BRCA-2.
Конверсия гена
Донором нового гена может быть:
гомологичная хромосома (во время мейоза)
систринская хроматида (так же во время мейоза)
дуплицированный ген той же хромосомы (во время митоза)
Исправление ошибок за счет 3’-5’ экзонуклеазной активности полимеразы при репликации (только у прокариот) (мутация E.coli mutD-мутатор-измен. -субъединицы ДНК-пол.III)
Тиминовые димеры, фермент фотолиаза ген-phr (у низших эукариот)
Удаление присоединенных алкильных и метильных групп - O-6-метилгуанинтрансфераза (ген ada)-удаляет О-6-метилгуанин
эксцизионная р. оснований [Е.coli ] [человек | узнавание поврежд. XPA-белком в ассоциации с RPA | привлекается ф-р транскр. TFIIH (P52, Р34, Р44, Р62, XPB-XPD-геликазн. акт-ть) | ERCC1-XPF, XPG – нуклеазы, надрезають ДНК по обе стороны от поврежд. | ДНК-полимераза- и вспомогат. белки RFC и PCNA застраивают брешь]
р. азотистых осн.-гликозилаза удаляет осн.
AP-сайт (апуриновый, апиримидиновый) | AP-эндонуклеаза опознает брешь, разрез. 5’-ДНК
пострепликативная р. (ПРР)
SOS-р. белки соед. с ДНК-полимеразой, доч. ДНК строится напротив поврежд. ДНК
Сокращения.
BER - Base Excision Repair
NER - Nucleotide Excision Repair
MMR - Mismatch Repair
NHEJ - Nonhomologous End-Joining
Мисмэтч репарация
В процессе реплакации, в результате ошибок полимеразы, могут встраиваться некомплиментарные нуклеотиды, что может привести к мутациям в дочерней цепи ДНК. Неспареные основания узнаются ферментами мисмэтч репарации и осуществляют замену ошибочно спаренных нуклеотидов.
Ферменты данной системы обеспечивают гомологичную рекомбинацию, а также задержку клеточного цикла в ответ на повреждения ДНК.
Система репарации ошибочно спаренных нуклеотидов у E. coli, использующая белки MutHLS, распознает и репарирует все некомплементарные пары оснований за исключением C–C. Кроме того, эта система репарирует небольшие вставки в одну из цепей ДНК, образующиеся в результате ошибок репликации, длина которых не превышает четырех нуклеотидов.
Обычно у E. coli ДНК метилирована Dam-метилазой
по сайтам GATC
. Однако после завершения репликации дочерняя цепь ДНК некоторое время остается неметилированной.
Эта система может быть реконструирована in vitro с использованием
ДНК с одной метилированной цепью в качестве субстрата, к которой добавляются очищенные белки MutH, MutL, MutS, UvrD (хеликаза II), холофермент ДНК-полимеразы III, ДНК-лигаза, белок SSB, а также одна из экзонуклеаз: ExoI, ExoVII или RecJ. Процесс репарации инициируется внесением одноцепочечного разрыва в неметилированную цепь вблизи частично метилированного сайта GATC с последующим гидролизом цепи ДНК и заполнением образующейся одноцепочечной бреши. При этом белок MutS связывается с ошибочно спаренными нуклеотидами. У белка MutL не обнаружено ферментативной активности, хотя он взаимодействует с MutS и необходим для активации MutH – эндонуклеазы, осуществляющей одноцепочечный разрыв ДНК. Таким образом, комплекс MutS–MutL, собранный на участке ДНК с ошибочно спаренным нуклеотидом, стимулирует эндонуклеазную (никазную) активность MutH. Бесклеточная система не требует присутствия MutH при наличии в ДНК-субстрате одноцепочечного разрыва. MutHLS-система репарации может
использовать частично метилированные последовательности GATC, расположенные выше и ниже поврежденного участка ДНК. При этом в
вырезании ошибочно включенного нуклеотида помимо хеликазы II принимает участие одна из экзонуклеаз: ExoI (3’-экзо), ExoVII (3’- и 5’-экзо) или RecJ (5’-экзо) в зависимости от расположения GATC-сайта по отношению к корректируемому нуклеотиду. Вслед за вырезанием нуклеотида образовавшаяся одноцепочечная брешь заполняется холоферментом ДНК-полимеразы III в присутствии SSB-белка и ДНК-лигазы. Следует подчеркнуть, что использование белка MutH и Dam-метилазы для распознавания дочерней цепи реплицировавшейся ДНК является уникальным свойством грамотрицательных бактерий. У грамположительных бактерий не происходит метилирование цепей ДНК в целях маркировки. Если сайты GATC полностью метилированы, MutHLS-система репарации E. coli изменяет ошибочно спаренные нуклеотиды в обеих цепях ДНК с одинаковой эффективностью.
У E. coli существуют, по крайней мере, еще два специфических
пути репарации ошибочно спаренных нуклеотидов. Система VSP (very short patchrepair pathway) репарирует некомплементарные пары G–T, заменяя их на G–C. Считается, что такие пары образуются в результате дезаминирования 5-метилцитозина в сайтах, где остатки С метилированы Dcm-метилазой. С более низкой эффективностью эта же система заменяет пары G–U на G–C. Другая MutY-зависимая система репарации специфически ликвидирует последствия окислительных повреждений гуанина. Если dGTP окисляется с образованием 8-оксо-dGTP, белок MutT расщепляет последний, предотвращая его включение в ДНК. Если же он все-таки включается напротив остатка С, то Fpg-гликозилаза (MutM) удаляет это модифицированное основание. В том случае, когда 8-оксо-G остается в составе ДНК, в следующем раунде репликации он спаривается с А, и в итоге может произойти трансверсия G–C>T–A. В этом случае белок MutY действует как ДНК-гликозилаза, удаляющая остаток A из некорректной пары, и как AP-лиаза, вносящая одноцепочечный
разрыв по соседству с AP-сайтом. Далее следуют процессы, уже рассмотренные выше в связи с функционированием системы репарации BER. Последовательность реакций с участием MutY также репарирует некомплементарные пары A–G и A–C с образованием соответственно пар C–G и G–C. Репарация ошибочно спаренных оснований у эукариот происходит при
участии комплекса белков, подобного системе MutHLS бактерий. Белок GTBP человека представляет собой гомолог бактериального белка MutS, а у дрожжей в соответствующей роли выступает белок Msh6. Распознавание ошибочно спаренных нуклеотидов у человека осуществляется гетеродимером MSH2–GTBP. Гомологами MutL в клетках S. cerevisiae являются белки MLH1 и PMS2, которые также существуют в виде гетеродимерных комплексов. Мутации в генах, кодирующих эти белки у человека, сопровождаются формированием мутаторного фенотипа и развитием наследственного неполипозного рака кишечника (синдром HNPCC – hereditary nonpolyposis colon cancer).
Прямая репарация
Существуют два основных пути репарации алкилированных оснований: эксцезионная репарация оснований (BER) и прямая репарация поврежденных оснований. При BER DNA-гликозилазы выщепляют цитотоксичные алкилированные основания в DNA на первом шаге с образованием AP-сайта и последующим его процессированием.В случае прямой репарации реализуются два способа: репарация алкилтрасферазами либо окисление алкильной группы, в обоих случаях происходит регенерация неповрежденных оснований. Если протекает репарация алкилтрасферазами (у млекопитающих только O 6 -алкилгуанин репарируется по этому пути), то O 6 -алкилгуанинтрансфераза (AGT) переносит метильную или этильную группу с O 6 -алкилгуанина на один из собственных остатков цистеина. Алкилированный в результате собственной активности белок инактивируется, но может служить регулятором активности своего гена и нескольких других. В отличие от суицидных O 6 -метилгуанинтрансфераз, направленно деметилирующих высокомутагенное и токсичное
повреждение O 6 -метилгуанин, AlkB из E. coli и его человеческие аналоги hABH2 и hABH3 окисляет метильные группы 1-метиладенина (1-meA) и 3-метилцитозина (3-meC) в DNA для регенерации немодифицированных оснований аденина и цитозина.
O 6 -алкилгуанинтрансфераза
O 6 -алкилгуанинтрансферазная активность обнаружена у большинства организмов и препятствует мутагенному действию O 6 -алкилгуанина. AGT превращает O 6 -алкилгуанин в гуанин, перемещая алкильную группу с DNA на реакционный остаток цистеина в белке в ходе необратимой реакции.
Такое ковалентное присоединение алкильной группы к остатку цистеина инактивирует фермент. Поэтому AGT - суицидный фермент, который подвергается протеолитической деградации после одной
реакции трансалкилирования. Структурные исследования позволяют обнаружить, что активный центр AGT расположен в объеме фермента на некотором расстоянии от DNA-связывающего участка. Считается, что фермент "работает" по механизму "переворачивания нуклеотида", чтобы плотно сблизить основание субстрата и нуклеофильный активный центр AGT.
Важность AGT в защите млекопитающих от токсического и мутагенного эффектов алкилирующих агентов была продемонстрирована на мышах. Трансгенные мыши с чрезмерной экспрессией AGT проявляют значительно более низкий уровень возникновения опухолей в ответ на действие метилирующего агента - N-метил-N-нитрозомочевины, в то время как мыши с дефицитом AGT оказывались на много более восприимчивы к инициированию опухоли и токсическим эффектам этого агента по сравнению с немутантными мышами. AGT - важный фермент в противоопухолевой терапии так как он препятствует цитотоксическим эффектам противоопухолевых агентов класса хлороэтилнитрозомочевины (CENU), например
BCNU (N,N-бис(2-хлороэтил)N-нитрозомочевина) или темозоломид. Было показано, что количество, в котором присутствует AGT в опухолях, в основном определяет на сколько благоприятным окажется исход противоопухолевой терапии с использованием CENU. CENU первоначально взаимодействует с O 6 -карбонильной группой гуанина с образованием соединения 4
, которое впоследствии преобразуется в N 1 ,O 6 -этаногуанин 5
. Соединение 5
перегруппируется в течение нескольких часов в физиологически активный ICL 6
. AGT препятствует образованию 6
при взаимодействии с 4
или 5
, возобновляя гуанин или формируя DNA-белковый аддукт 8
. Экспериментально получено подтверждение образования аддукта 8
, но он был выделен в слишком малом количестве для его детального описания. При биохимическом исследовании этой проблемы был введен N 1 ,O 6 -этаноксантин
9
как стабильный аналог 5
в DNA. Этаноксантин 9
реагирует с AGT с образованием стабильного DNA-белкового аддукта 7
. Этот подход позволил формировать ковалентно связанный AGT-DNA аддукт 10
в большом количестве, который может быть использован для определения структуры AGT связанного с DNA. Взаимовлияние AGT и алкилирующей терапии привело к поиску ингибиторов AGT, которые могут быть использованы в терапии рака в сопряжении с алкилирующими агентами. К настоящему времени созданы ингибиторы, главным образом, производные гуанина с заместителями в O 6 -позиции. O 6 -бензилгуанин 2
был обнаружен как типичный ингибитор AGT, с которым сравнивают новые молекулы. Эффективность O 6 -BzG в усилении цитотоксичности CENU продемонстрирована на моделях животных. Лимитирующим фактором этого терапевтического подхода является токсичность для здоровых органов, частично для костного мозга. Некоторые
группы ученых направлены обойти эту проблему путем генерирования ATG-варианта, устойчивого к ингибированию O 6 -BzG, что может быть использовано для защиты костного мозга при помощи генного переноса.

Оксидоредуктазы AlkB, hABH2 и hABH3
Еще один путь прямой репарации алкилированных оснований - окисление алкильной группы с регенерацией неповрежденного азотистого основания. Фермент AlkB у Escherichia coli и два человеческих аналога hABH2 и hABH3 направленно деметилируют 1-метиладенин и 3-метилцитозин в DNA. Но в отличие от AGT, эти ферменты обладают субстратной специфичностью, направленной на поверхность пар оснований G:C и A:T. Повреждения 1-алкиладенин
и 3-метилцитозин формируются когда аденин и цитозин находятся в одноцепочечной структуре (в период репликации или транскрипции) и являются субстратами для AlkB, hABH2 и hABH3. Они окисляют метильные группы 1-метиладенина (1-meA) и 3-метилцитозина (3-meC) в DNA для регенерации азотистых оснований аденина и цитозина. Было также показано, что AlkB защищает от токсичного повреждения - аддукта с этильной группой и преобразует 1-этиладенин в аденин в DNA, продуцирующий в результате реакции ацетальдегид. Таким образом репарируются повреждения известного мутагена и канцерогена этиленоксида, эндогенно образующегося в ходе метаболизма этилена, а также широко применяющегося как фумигант для стерилизации. Аддукты с гидроксиэтилом, генерируемые этиленоксидом обнаружены в DNA клетки. Другие малые алкилирующие эпоксиды также задействованы в большом количестве в химическом производстве. Было показано, что AlkB снижает токсические эффекты DNA-повреждающих агентов, генерирующих гидроксиэтильный,
пропильный и гидроксипропильный аддукты. AlkB репарирует алкилированные трифосфаты 1-me-dATP активно, но неэффективно. Предполагалось, что эта способность может снижать уровень встраивания алкилированных трифосфатов при синтезе DNA; к тому же Фрагмент Кленова DNA полимеразы I у E. coli может использовать 1-me-dATP как предшественник для синтеза DNA in vitro. Человеческие ферменты hABH2 и hABH3 также деметилировали 1-метиладениновые остатки в поли(dA), они были неэффективны на коротких субстратах. Таким образом hABH3 обладал очень низкой активностью на тримере d(Tp1-meApT), тогда как у hABH2 активности не было обнаружено.
AlkB и его человеческие аналоги является частью -кетоглутарат/Fe(II)-зависимого суперсемейства диоксигеназ, и в процессе репарации протекает свместно декарбоксилирование -кетоглутарата и окислительное деметилирование поврежденного основания. Повреждения 1-meA и 3-meC формируются в основном в одноцепочечной DNA и предположительно
возникают в репликативной вилке и в активно транскрибируемых генах где они могут заблокировать DNA- и RNA-полимеразы. В действительности AlkB, hABH2 и hABH3 репарируют эти повреждения в одноцепочечной DNA, но также протекает репарация олигонуклеотидов, отожженных с комплементарной цепью после алкилирования. Эффективность репарации 1-метиладенина AlkB не зависит от полинуклеотидной структуры, но необходимо наличие нуклеотид-5"-фосфатной группы. Также человеческие ферменты hABH2 и hABH3 деметилировали остатки 1-метиладенина в поли(dA), они были неэффективны на коротких субстратах. К тому же, повреждения, содержащие положительный заряд (рибонуклеозиды 1-meA и 3-meC имеют соответственно pKa= 9.3 и 9.6) лучше репарировались, чем незаряженные основания (1-meG и 3-meT). Однако, так как 1-meG (и в меньшей степени 3-meT) репарировались AlkB, то формальный положительный заряд основания не является необходимым условием для функционирования AlkB. С этой точки зрения
пока неясно, зависит ли этот результат от того, что положительно заряженные основания лучше распознаются посредством электростатических взаимодействий с AlkB или положительно заряженные основания просто делают DNA лучшей уходящей группой после гидроксилирования метильной группы.
Сокращения:
- AGT - алкилгуанин трансфераза
- BER - эксцезионная репарация оснований
- DNA - дезоксирибонуклеиновая кислота
- RNA - рибонуклеиновая кислота
- BCNU - N,N-бис(2-хлороэтил)N-нитрозомочевина
- CENU - хлороэтилнитрозомочевина
- O 6 -BzG - O 6 -бензилгуанин
- 1-meA - 1-метиладенин
- 1-meG - 1-метилгуанин
- 3-meC - 3-метилцитозин
- 3-meT - 3-метилтимин
Литература:
» Orlando D. Scharer (2003) Angew. Chem. Int. Ed. 42, 2946-2974
» James C. Delaney and John M. Essigmann Mutagenesis, genotoxicity, and repair of 1-methyladenine, 3-alkylcytosines, 1-methylguanine, and 3-methylthymine in alkB Escherichia coli
» Pertti Koivisto, Tod Duncan, Tomas Lindahl, and Barbara Sedgwick Minimal Methylated Substrate and Extended Substrate Range of Escherichia coli AlkB Protein, a 1-Methyladenine-DNA Dioxygenase*
» Duncan, T., Trewick, S. C., Koivisto, P., Bates, P. A., Lindahl, T. & Sedgwick, B. (2002) Proc. Natl. Acad. Sci. USA 99, 16660-16665. 5. Aas, P. A., Otterlei, M., Falnes, P. O., Vagbo, C. B., Skorpen, F., Akbari, M., Sundheim, O., Bjoras, M., Slupphaug, G., Seeberg, E., et al. (2003) Nature 421, 859-863.
» Hollis, T., Lau, A., and Ellenberger, T. (2000) Mutat. Res. 460, 201-210
» Daniels, D. S., and Tainer, J. A. (2000) Mutat. Res. 460, 151-163
» Trewick, S. C., Henshaw, T. F., Hausinger, R. P., Lindahl, T., and Sedgwick, B. (2002) Nature 419, 174-178
» Falnes, P. O., Johansen, R. F., and Seeberg, E. (2002) Nature 419, 178-181
» Duncan, T., Trewick, S. C., Koivisto, P., Bates, P. A., Lindahl, T., and Sedgwick, B. (2002) Proc. Natl. Acad. Sci. U. S. A. 99, 16660-16665
» Aas, P. A., Otterlei, M., Falnes, P. O., Vagbo, C. B., Skorpen, F., Akbari, M., Sundheim, O., Bjoras, M., Slupphaug, G., Seeberg, E., and Krokan, H. E. (2003) Nature 421, 859-863
» Aravind, L., and Koonin, E. V. (2001) Genome Biology 2, 0007.1-0007.8
» Bodell, W. J., and Singer, B. (1979) Biochemistry 18, 2860-2863
» Boiteux, S., and Laval, J. (1982) Biochimie (Paris) 64, 637-641
» Larson, K., Sahm, J., Shenkar, R., and Strauss, B. (1985) Mutat. Res. 150, 77-84
» Dinglay, S., Trewick, S. C., Lindahl, T., and Sedgwick, B. (2000) Genes Dev. 14, 2097-2105
Репарация разрывов ДНК
Фотореактивация
Абсорбция энергии УФ излучения молекулами ДНК приводит к образованию различных типов повреждений. Хотя одно- и двуцепочечные разрывы, а также ДНК-белковые сшивки могут возникать, большая часть индуцированных УФ-излучением повреждений приходится на модификацию азотистых оснований, с образованием циклобутан пиримидиновых димеров (CPD) и пиримидин-пиримидоновых фотопродуктов (6-4PP), как наиболее часто встречающихся типов фотоповреждений.
Пиримидиновые димеры являются ингибиторами и для репликации, и для транскрипции, что замедляет рост и приводит к мутагенезу в процессе репликации ДНК, если такие повреждения остаются неотрепарированными.
Для удаления индуцированных светом повреждений в ДНК у многих организмов применяются ферменты, специфично связывающиеся с CPD (CPD-фотолиаза) или с 6-4PP (6-4PP-фотолиазы) и исправляющие эти повреждения. CPD-фотолиазы обнаружены в бактериях, грибах, растениях, беспозвоночных и во многих позвоночных, 6-4PP-фотолиазы обнаружены, пока, только в Drosophila, тутовом шелкопряде, Xenopus laevis и в гремучих змеях, но не у Escherichia coli или дрожжей. У людей не обнаружено фотолиаз. Фотолиазы содержат FAD как каталитический кофактор и дополнительный хромофор в качестве светособирающей антенны.
Дополнительные хромофоры – это или 5,10-метенилтетрагидрофолат (MTHF), или 8-гидрокси-5-деазорибофлавин (8-HDF), с максимумами поглощений на длинах волн 380 и 440 нм, соответственно. Кристаллические структуры CPD-фотолиаз E.coli и Anacystis nidulans подтверждают, что для связывания с ДНК ферменты поворачивают пиримидиновый димер из дуплекса в углубление, содержащее каталитический кофактор. Затем циклобутановое кольцо расщепляется в ходе переноса, индуцированного светом электрона. CPD-фотолиазы селективно распознают CPD, подобно ДНК-связывающим белкам. Белый свет или UV-B-излучение индуцируют экспрессию CPD-фотолиаз. В отличие от CPD-фотолиаз, 6-4PP-фотолиаза стабильно экспрессируется и не регулируется ни белым светом, ни UV-B-излучением.
Схематичное изображение фотореактивации в хроматине. Октамеры гитонов голубого цвета, ДНК - черного. Фотолиаза связывается с циклобутан-пиримидиновыми димерами (CPD), поворачивает пиримидиновый димер и регенерирует природные пиримидины в ходе, зависимой от освещения, реакции. Фотолиаза предпочтительно репарирует CPD в линкетной ДНК. Репарация в нуклеосомах замедлена и, предположительно, облегчается динамическими свойствами нуклеосом, перемещающих повреждения ДНК в область линкерной ДНК.
Класс CDP-специфичных фотолиаз из микроорганизмов, определенный как класс I фотолиаз, был первым охарактеризованным представителем семейства фотолиаз. Близкородственный класс фотолиаз специфичных к 6-4-фотопродуктам был обнаружен недавно, представители этого семейства были найдены в Drosophila melanogaster, Xenopus laevis и Arabidopsis thaliana. Криптохромы, являющиеся фоторецепторами на фиолетовую часть светового спектра обнаруженные в растениях и других организмах, также являются близкородственными классу I фотолиаз.
Более дальнородственным семейством CPD-фотолиаз, названное классом II фотолиаз было идентифицировано в ряде видов, включая животных, Archaebacterium, Eubacterium и высших растений. Было показано, что все охарактеризованные фотолиазы содержат восстановленный FAD и большинство содержат вторичные хромофоры в зависимости от вида либо MTHF, либо 8-HDF. Подобный механизм реакции был предложен для обоих классов CPD-фотолиаз. Хромофор MTHF или 8-HDF подобен антенне, которая абсорбирует фиолетовый свет и расходует поглощенную энергию на регенерацию FADH-.
Состав кофактора растительной фотолиазы не до конца исследован, хотя, недавно было показано, что CPD-фотолиазы из Arabidopsis содержавшие только FADH обладали ферментативной активностью.
Литература:
» Fritz Thoma, Light and dark in chromatin repair: repair of UV-induced DNA lesions by photolyase and nucleotide excision repair, Institut fur Zellbiologie, ETH-Zurich, Honggerberg, CH-8093 Zurich, Switzerland
» Characterization of Arabidopsis photolyase enzymes and analysis of their role in protection from ultraviolet-B radiation, Wanda M. Waterworth 1, Qing Jiang, Christopher E. West, M. Nikaido and Clifford M. Bray
Повреждённых при нормальном биосинтезе ДНК в клетке или в результате воздействия физических или химических реагентов. Осуществляется специальными ферментными системами клетки. Ряд наследственных болезней (напр., пигментная ксеродерма) связан с нарушениями систем репарации.
История открытия
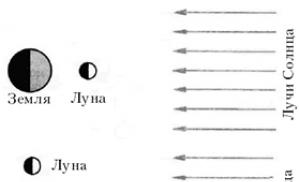
Начало изучению репарации было положено работами Альберта Кельнера (США), который в 1948 году обнаружил явление фотореактивации - уменьшение повреждения биологических объектов, вызываемого ультрафиолетовыми (УФ) лучами, при последующем воздействии ярким видимым светом (световая репарация ).
Р. Сетлоу, К. Руперт (США) и другие вскоре установили, что фотореактивация - фотохимический процесс, протекающий с участием специального фермента и приводящий к расщеплению димеров тимина , образовавшихся в ДНК при поглощении УФ-кванта.
Позднее при изучении генетического контроля чувствительности бактерий к УФ-свету и ионизирующим излучениям была обнаружена темновая репарация - свойство клеток ликвидировать повреждения в ДНК без участия видимого света. Механизм темновой репарации облучённых УФ-светом бактериальных клеток был предсказан А. П. Говард-Фландерсом и экспериментально подтверждён в 1964 году Ф. Ханавальтом и Д. Петиджоном (США). Было показано, что у бактерий после облучения происходит вырезание повреждённых участков ДНК с изменёнными нуклеотидами и ресинтез ДНК в образовавшихся пробелах.
Системы репарации существуют не только у микроорганизмов , но также в клетках животных и человека , у которых они изучаются на культурах тканей . Известен наследственный недуг человека - пигментная ксеродерма , при котором нарушена репарация.
Источники повреждения ДНК
Основные типы повреждения ДНК
Устройство системы репарации
Каждая из систем репарации включает следующие компоненты:
- ДНК-хеликаза - фермент, «узнающий» химически изменённые участки в цепи и осуществляющий разрыв цепи вблизи от повреждения;
- ДНКаза (дезоксирибонуклеаза) - фермент, "разрезающий" 1 цепочку ДНК (последовательность нуклеотидов) по фосфодиэфирной связи и удаляющий повреждённый участок: экзонуклеаза работает на концевые нуклеотиды 3` или 5`, эндонуклеаза - на нуклеотиды, отличные от концевых;
- ДНК-полимераза - фермент, синтезирующий соответствующий участок цепи ДНК взамен удалённого;
- ДНК-лигаза - фермент, замыкающий последнюю связь в полимерной цепи и тем самым восстанавливающий её непрерывность.
Типы репарации
Прямая репарация
Прямая репарация - наиболее простой путь устранения повреждений в ДНК, в котором обычно задействованы специфические ферменты , способные быстро (как правило, в одну стадию) устраняют соответствующее повреждение, восстанавливая исходную структуру нуклеотидов . Так действует, например, O6-метилгуанин-ДНК-метилтрансфераза , которая снимает метильную группу с азотистого основания на один из собственных остатков цистеина .
Эксцизионная репарация
Эксцизионная репарация (англ. excision - вырезание) включает удаление повреждённых азотистых оснований из ДНК и последующее восстановление нормальной структуры молекулы по комплементарной цепи. Ферментативная система удаляет короткую однонитевую последовательность двунитевой ДНК, содержащей ошибочно спаренные или поврежденные основания, и замещает их путём синтеза последовательности, комплементарной оставшейся нити.
Эксцизионная репарация является наиболее распространённым способом репарации модифицированных оснований ДНК . Она базируется на распознавании модифицированного основания различными гликозилазами, расщепляющими N-гликозидную связь этого основания с сахарофосфатным остовом молекулы ДНК. При этом существуют гликозилазы, специфически распознающие присутствие в ДНК определенных модифицированных оснований (оксиметилурацила, гипоксантина, 5-метилурацила, 3-метиладенина, 7-метилгуанина и т. д.). Для многих гликозилаз к настоящему времени описан полиморфизм, связанный с заменой одного из нуклеотидов в кодирующей последовательности гена. Для ряда изоформ этих ферментов была установлена ассоциация с повышенным риском возникновения онкологических заболеваний [ Chen, 2003 ].